|
|
 |
|||||||||||||||||||||||||||||||
|
|
باکتری شناسی | ایمنی شناسی | قارچ شناسی | انگل شناسی | ویروس شناسی | |||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
|
واژه های کلیدی
گلایکولیز
|
از جمله نیازهای باکتری برای رشد می توان به منبع انرژی ،منبع کربن آلی(مثل قندها و اسیدهای چرب) و یونهای فلزی(مثل آهن) اشاره کرد.دمای مناسب،pH ونیاز(و یا عدم نیاز) به اکسیژن نیز برای رشد باکتری مهم هستند.
نیاز به اکسیژن باکتریهای هوازی اجباری باید در حضور اکسیژن رشد کنند و توان تخمیر کردن را ندارند.اما باکتریهای بیهوازی اجباری،فسفوریلاسیون اکسیداتیو را انجام نمی دهند.علاوه براین،آنها در حضور اکسیژن کشته می شوند.این باکتریها فاقد آنزیمهای خاصی مثل کاتالاز(که پراکسید هیدروژن را به آب و اکسیژن تجزیه می کند)،پرکسیداز(که بواسطه آن NADH + H2O2 به NAD و O2 تبدیل می شوند) و سوپراکسید دیسموتاز(که بواسطه آن سوپراکسید به پراکسید هیدروژن تبدیل می شود) هستند.این آنزیمها سمیت پراکسید و رادیکالهای آزاد اکسیژن را که طی متابولیسم در حضور اکسیژن تولید شده اند برطرف می کنند. ،باکتریهایی هستند که به شکل بیهوازی تنفس دارند ولی می توانند در حضور اکسیژن هم زنده بمانند. باکتریهای بیهوازی مقاوم به هوا(آئروتولرنت) باکتریهای بیهوازی اختیاری می توانند هم تخمیر و هم تنفس هوازی را انجام دهند.به طور کلی در حضور اکسیژن،تنفس بیهوازی غیر فعال می شود و این ارگانیسمها به شکل هوازی تنفس می کنند. باکتریهای میکروآئروفیل،به خوبی در غلظتهای کم اکسیژن رشد می کنند ولی در غلظتهای بالاتر کشته می شوند.
نیازهای غذایی عبارتند از منابع کربن آلی،نیتروژن،فسفر،گوگرد و یونهای فلزی از جمله آهن.باکتریها مولکولهی کوچکی ترشح می کنند که به آهن متصل می شوند(سایدروفورها مثل انتروباکتین و مایکوباکتین). سپس سایدروفورها(به همراه آهن متصل به آنها) از طریق گیرنده هایی وارد سلول باکتری می شوند.انسان میزبان باکتری هم دارای پروتئینهای انتقالی آهن می باشد(مثل ترانسفرین).بنابراین،باکتریهایی که رقابت ناکارامدی برای کسب آهن با میزبان دارند پاتوژنهای ضعیفی هستند. دما باکتریها ممکن است در طیف وسیع دمایی از دمای نزدیک انجماد گرفته تا دمای نزدیک نقطه جوش آب رشد کنند.آنهایی که بهترین رشد را در میانه این محدوده دارند مزوفیل محسوب می شوند که همه پاتوژنهای انسانی و باکتریهای فرصت طلب جزو این گروه هستند.(باکتریهایی که دمای رشد مناسب بالاتر و پایین تری دارند به ترتیب ترموفیل و سایکروفیل نامیده می شوند).
pH بسیاری از باکتریها بهترین رشد را در pH خنثی دارند،اگرچه باکتریهای خاصی هم وجود دارند که توان بقا و رشد در شرایط اسیدی یا قلیایی را دارند
|
|||||||||||||||||||||||||||||||
|
باکتریها و تقسیم دوتایی
|
||||||||||||||||||||||||||||||||
|
|
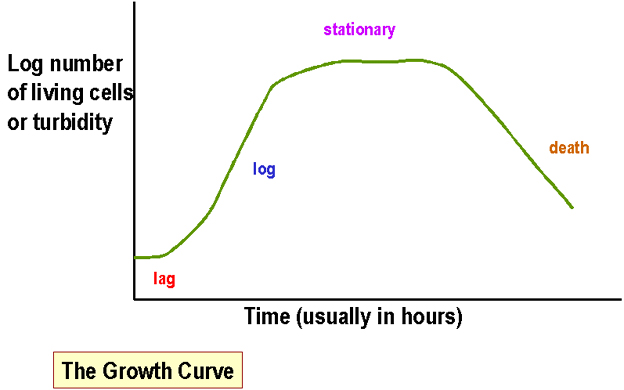
اندازه گیری جرم باکتریها در کشت مایع باکتریها روشهای معمول عبارتند از: الف)کدورت - (کدورت کشت مایع باکتریها نشاندهنده کل آنها(زنده و مرده) می باشد.این ویژگی معمولا" بوسیله اسپکتروفتومتر اندازه گیری می شود).شکل 1
در هر دو روش،ترسیم لگاریتم کدورت یا تعداد سلولهای زنده در مقابل زمان به عنوان "منحنی رشد"مطرح می شود.زمان تقسیم به عنوان زمان لازم برای دوبرابر شدن توده باکتریایی تعریف می شود.
|
|||||||||||||||||||||||||||||||
|
منابع اینترنتی
چرخه
کربس
|
متابولیسم قندها (به عنوان مثالی ازمسیرهای متابولیک) گلایکولیز (مسیر امبدن،مایرهوف،پارناز [EMP] ) این مسیر،رایج ترین مسیر کاتابولیسم قندها در باکتریهاست(این مسیر در اکثر سلولهای گیاهی و جانوری نیز یافت شده).یک سری فرایندهای آنزیمی منجر به تبدیل قندها به پایرووات،تولید ATP و NADH می شود.انرژی شیمیایی لازم برای بیوسنتز در ترکیباتی که شکل می گیرند ذخیره می شود (NADH و ATP)
* به تعداد کربن در مولکول اشاره می کند
مسیرهای جایگزینی برای این مسیر جهت کاتابولیزه کردن قندها برای تولید انرژی ذخیره شده در ATP وجود دارد. این مسیرها شامل مسیر پنتوز-فسفات(شانت هگزوز منوفسفات) می باشد که در اکثر سلولهای گیاهی و جانوری وجود دارد.NADPH بوسیله این مسیر تولید می شود.مسیر دیگر،مسیر انتنر دودوروف می باشد که به طور کلی فقط در باکتریهای خاصی دیده می شود.
تنفس بیهوازی تنفس بیهوازی شامل گلایکولیز و تخمیر می باشد.در طی مرحل پایانی این فرایند،NADH (محصول گلایکولیز) با از دست دادن یک هیدروژن به NAD تبدیل می شود.این هیدروژن به پایرووات اضافه می شود و بسته به نوع گونه باکتری،طیف وسیعی از محصولات نهایی تولید می شوند.
تنفس هوازی تنفس هوازی شامل گلایکولیز و چرخه کربس(TCA) می باشد.پایرووات کاملا" به دی اکسید کربن شکسته می شود و در طی این فرایند،NAD به NADHتبدیل می شود. بنابراین،در تخمیر هوازی،NADH از دو منبع تولید می شود(گلایکولیز و چرخه کربس).در فسفوریلاسیون اکسیداتیو،NADH اضافی مجددا" به NAD تبدیل می شود و در طی آن ATP بیشتری تولید می شود. یوبی کوینونها و سایتوکرومها،اجزای زنجیره انتقال الکترون هستند که در این فرایند شرکت دارند.تبدیل اکسیژن به آب،آخرین مرحله این فرایند است. چرخه کربس(ترکیبات حدواسط 4 تا 6 کربنه)
|
|||||||||||||||||||||||||||||||
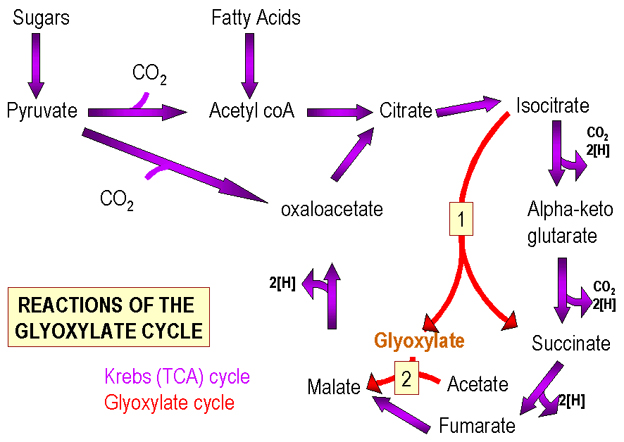
شکل2.چرخه های کربس و گلی اکسالات |
فسفوریلاسیون اکسیداتیو
چرخه کربس(شکل 2) حاوی مولولهای حدواسط 6-4 کربنه است.پایرووات (C3)می تواند طوری به چرخه کربس افزوده شود که تعداد این ترکیبات ثابت بماند یا افزایش پیدا کند. 1)حذف CO2 (C1) از پایرووات و تبدیل به استیل کوآنزیمA وبه دنبال آن اضافه شدن به یک جزء C4چرخه(اگزالواستات)و تولید یک جزء C6(اسید سیتریک).بنابراین،تعداد مولکولهای شش کربنه حاصل،با تعداد مولکولهای چهار کربنه اولیه برابر است.
2)با اضافه شدن CO2 به پایرووات،یک ترکیب 4 کربنه تولید می شود.در این مثال،مولکولهای 4 کربنه شکل می گیرند:
بنابراین اگر بعضی اجزای چرخه برای مصرف در دیگر مسیرهای بیوسنتتیک از چرخه حذف شوند می توانند از طریق این واکنش مجددا" ساخته شوند.
|
|||||||||||||||||||||||||||||||
|
منابع اینترنتی |
||||||||||||||||||||||||||||||||
|
(زنجیره
زوج)
|
متابولیسم اسیدهای چرب اسیدهای چرب به گروههای استیل شکسته می شوند (C2) که وارد چرخه کربس شده و به یک مولکول حد واسط چهار کربنه اضافه شده و محصول شش کربنه حاصل می شود.در طول چرخه، C2به شکل CO2 حذف شده و C4مجددا" شکل می گیرد. درمجموع،تغییری در تعداد مولکولهایحدواسط چرخه رخ نمی دهد:
به جای این چرخه،باکتریها از چرخه گلی اکسالات(شکل2) استفاده می کنند که در واقع نوعی چرخه کربس تغییر یافته است که در آن مراحل آنزیمی که دو مولکول CO2 از واسط شش کربنی حذف شده اند میانبر زده شده اند. واسط شش کربنی به دو ترکیب چهار کربنی تبدیل می شود.بنابراین،ممکن است برای هر گروه استیل(از اسیدهای چرب)،یک مولکول واسط در چرخه بوجود بیاید.مسیر گلی اکسالات معمولا" به دلیل اینکه سلولهای حیوانی از اسید چرب پیش ساخته در غذا استفاده می کنند در آنها دیده نمی شود.
به طور خلاصه،فعالیت چرخه کربس به شکل بیوسنتز و تولید انرژی می باشد.هر چند که واسطه های این چرخه برای مصرف در دیگر چرخه های متابولیک از چرخه خارج می شوند ولی باید مجددا" تامین شوند که این فرایند برای بکرگیری قندها و اسیدهای چرب متفاوت است.
|
|||||||||||||||||||||||||||||||
|
|
|
|||||||||||||||||||||||||||||||