|
|
 |
||||
|
|
PARASITOLOGÍA |
||||
|
|||||
|
|
|||||
|
E-MAIL |
|||||
|
|
|||||
|
PALABRAS CLAVE |
En esta sección se discutirá la estructura de las envolturas celulares de Gram negativos, Gram positivos y ácido alcohol resistentes. También se describirá la composición y la función de las macromoléculas únicas de la envoltura celular y se mencionará su biosíntesis. Además, se abordarán las endosporas, que son de muchas maneras estructuras bacterianas inusuales, (incluyendo la estructura de su envoltura ). Envoltura celular La envoltura celular puede definirse como: la membrana celular y la pared celular, mas la membrana externa si ésta se encuentra presente. La pared celular consiste de la capa de peptidoglicano y otras estructuras que ésta lleva unidas. La mayoría de las envolturas celulares caen en dos categorías principales (Figura 1): Gram positivas y Gram negativas. Esto se basa en las características de la tinción de Gram en donde se reflejan grandes diferencias entre los dos grupos. Otros tipos de pared celular se encuentran en pocas especies bacterianas (ni en los Gram positivos ni en los Gram negativos). El peptidoglicano (PG) es una macromolécula única, con enlaces altamente entrecruzados, forma una bolsa que rodea a la membrana celular bacteriana y que concede rigidez a la envoltura. En tamaño molecular es inmensa (su peso molecular es del orden de millones; comparado con las proteínas que son del orden de miles en peso molecular). El peptidoglicano consiste de: un esqueleto de glicano (polisacárido) que está compuesto de ácido N-acetil murámico y N-acetil glucosalina, cadenas laterales de péptidos conteniendo D- y L- aminoácidos y algunas veces ácido diaminopimélico. Las cadenas laterales están unidas de manera entrecruzada por puentes peptídicos. La estructura de estos puentes peptídicos varían entre las especies bacterianas. El ácido murámico, los D-aminoácidos y el ácido diaminopimélico no son sintetizados por mamíferos. El PG se encuentra en todas las eubacterias excepto en Chlamydia o en Mycoplasma.
|
||||
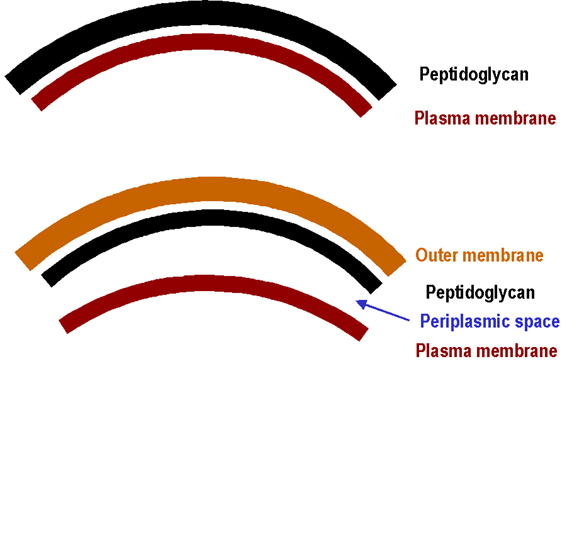 Figure 1.
Diagrama simplificado de la envoltura celular de bacterias Gram positivas y
Gram negativas
Figure 1.
Diagrama simplificado de la envoltura celular de bacterias Gram positivas y
Gram negativas |
Envoltura de las Gram Positivas (Figura 2) Unidos covalentemente a la gruesa capa de peptidoglicano están los ácidos teicóicos (sus esqueletos son usualmente polímeros del ribitol o del glicerol que contienen fósforo) o el ácido teicurónico (polisacáridos que contienen ácido glucurónico). Estas moléculas, cargadas negativamente se cree que están involucradas en la concentración de iones metálicos de los alrededores. Los ácidos teicóicos pueden también dirigir a las enzimas autolíticas hacia los sitios de digestión del peptidogicano (autolisis), uno de los pasos en la biosíntesis de la pared celular. Algunas veces también están presentes polisacáridos neutros. El ácido lipoteicóico en muchas bacterias se asocia generalmente con la membrana celular. Otras veces ocurre en las fimbrias, en el exterior de la célula, expresado en el exterior puede estar involucrado en la adhesión o adherencia a las células epiteliales, permitiendo así que se lleve a cabo la colonización, digamos de la garganta (ejemplo: en el Streptococcus del Grupo A).
|
||||
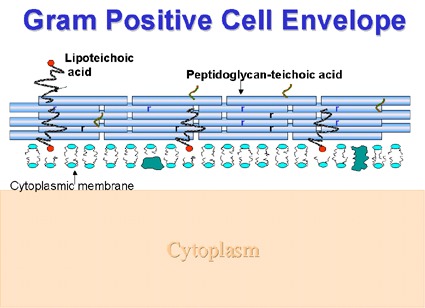 Figura
2.
Los
componentes estructurales de las envolturas celulares de los Gram positivos Figura
2.
Los
componentes estructurales de las envolturas celulares de los Gram positivos
|
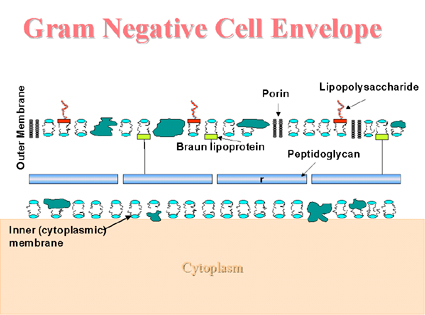
La envoltura celular de los Gram negativos (Figura 3) Unida covalentemente a la delgada capa de peptidoglicano está la lipoproteína de Braun, la cual une a la membrana externa con la pared celular. Como otras membranas, la membrana externa contiene fosfolípidos y proteínas, pero a diferencia de las otras membranas, ésta contiene moléculas adicionales como el lipopolisacárido (LPS). El lipopolisacárido es importante para la pared celular ya que le ayuda a proveer una barrera de permeabilidad ante substancias hidrofóbicas. El LPS consiste de tres regiones: el antígeno O, de una región intermedia o central y la región mas interna que es el lípido A. La región central contiene varios azúcares (heptosas u ácido ceto-desoxi-octónico), que no se encuentran en ninguna otra estructura en la naturaleza y el lípido A contiene ácidos grasos tipo β hydroxi (poco comunes en la naturaleza). Esta molécula despliega una actividad endotóxica, es decir es una endotoxina. Las porinas en la membrana externa forman los canales que permitirán el paso a los pequeños nutrientes hidrofílicos (tales como los azúcares) a través de la membrana externa. Bacterias ácido alcohol resistentes y bacterias relacionadas (mycobacteria, nocardia y corynebacteria). Las envolturas celulares de estos organismos son considerablemente mas complejos que otras bacterias. Los ácidos micólicos (ácidos grasos largos y remificados) están covalentemente unidos al peptidoglicano, vía un polisacárido. Otros compuestos conteniendo ácido micólico y más lípidos complejos forman una capa laminada, membranosa y cerosa por fuera de la capa de peptidoglicano. |
||||
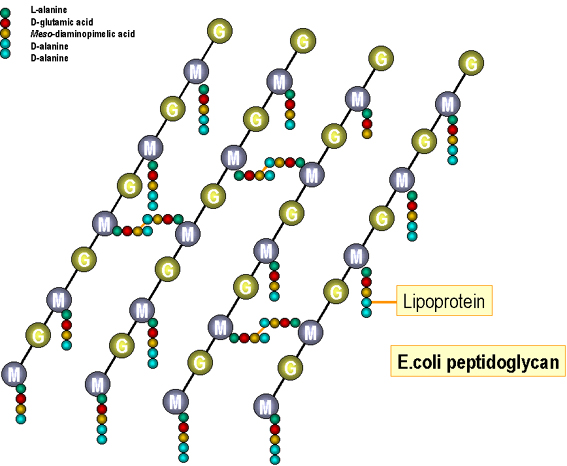 Figura 4.
Estructure del peptidoglicano de E. coli
Figura 4.
Estructure del peptidoglicano de E. coli
|
Síntesis de las macromoléculas de la envoltura cellular. El peptidoglicano (Figuras 4 y 5): La subunidad precursora (el muramil pentapeptido unido a uridina difosfato, UDP) se sintetiza en el citoplasma y atraviesa la membrana celular. La subunidad es enzimáticamente movilizada desde el nucleótido hasta el lípido acarreador (undecaprenol/bactoprenol) y ensamblado en una subunidad completa (pentapéptido disacárido unido a un puente peptídico). Las subunidades completas entonces son exportadas hacia la pared celular. Después de haberse liberado el monómero de undecaprenol, este es recirculado a la membrana celular y puede ser usado de nuevo. Los esqueletos de glicano de la pared celular existente, se rompen enzimaticamente (por autolisinas) para permitir la inserción de subunidades sintetizadas de novo. Si estas enzimas están sobre-activadas, la pared celular resulta degradada y la presión osmótica alta de la célula revienta a la membrana citoplásmica, produciendo la muerte de la célula (“autolisis”). Las uniones por entrecruzamiento de las cadenas peptídicas laterales de la subunidad de peptidoglicano insertada a la cadena existente se llevan a cabo enzimaticamente (intervienen proteínas de unión a penicilina). Las subunidades completas de los ácidos teicóicos y teicurónicos también se sintetizan en la membrana celular (en acarreadores lipídicos) antes de su transporte e inserción a la pared celular pre-existente.
|
||||
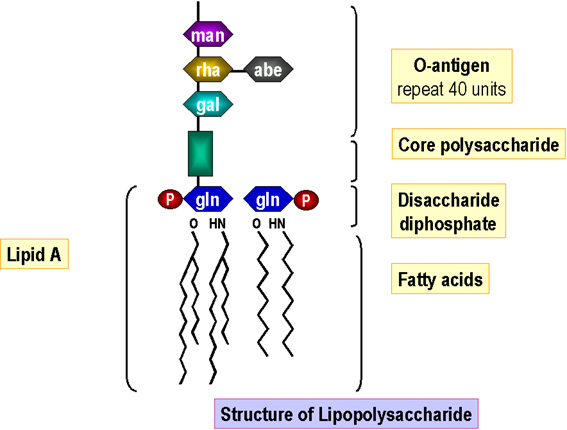 Figura 6. Estructura del lipopolisacárido
Figura 6. Estructura del lipopolisacárido |
Lipopolisacárido El ensamblaje del lípido A se lleva a cabo en la membrana celular y los azúcares centrales se le van uniendo secuencialmente. Las subunidades del antígeno-O son sintetizadas independientemente (sobre un lípido acarredor, como en la síntesis del peptidoglicano). El antígeno-O completamente sintetizado se une entonces a la región central y el lípido A, generándose el lipopolisacárido) en la membrana celular, antes de su paso/inserción a la membrana externa (Figura 6).
|
||||
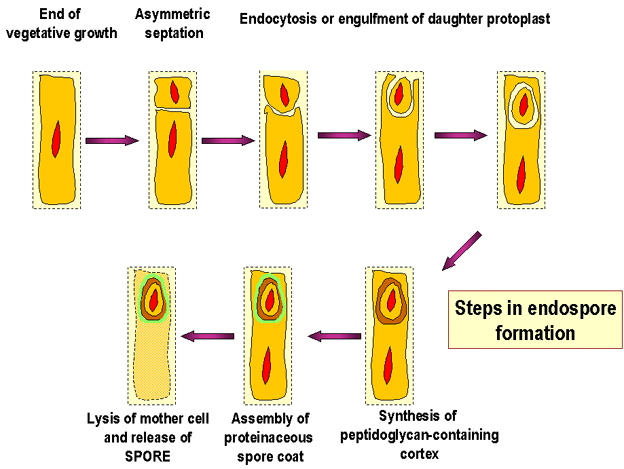 Figura 7. Pasos en la formación de la endospora
Figura 7. Pasos en la formación de la endospora
|
Endosporas Estas células bacterianas, Gram positivas modificadas, tienen una envoltura celular inusual que consta de una membrana celular y una membrana externa. La capa de peptidoglicano es menos entrecruzada que en la mayoría de las células bacterianas y contiene una forma deshidratada de ácido murámico. El peptidoglicano de la espora se mencionado como la corteza y se encuentra entre las dos membranas. Es una cubierta consistente en queratina altamente entrecruzada que se encuentra rodeando el exterior de la célula. Debido a esta cubierta la espora bacteriana es altamente resistente a los agentes químicos. En la replicación bacteriana, normalmente las células se dividen, se forma un septo separando a la célula madre en dos hijas de tamaño más o menos equivalente. Cuando ocurre la esporulación, la division celular no es equitativa y la célula más grande, llamada “célula madre” envuelve a la “célula hija”. La membrana celular de la célula hija constituye la membrana interna de la espora y la membrana celular de la célula madre forma la membrana externa (Figuras 7 y 8).
|
||||
|
PELÍCULA |
|||||
|
|
|
||||