|
x |
x |
|
 |
 |
|
BỆNH NHIỄM TRÙNG |
VI KHUẨN HỌC |
MIỄN DỊCH HỌC |
NẤM HỌC |
KÝ SINH TRÙNG HỌC |
VIRÚT HỌC |
|
ENGLISH |
MIỄN DỊCH HỌC – CHƯƠNG
MƯỜI
PHỨC HỢP HÒA HỢP MÔ CHỦ
YẾU (MHC) VÀ THỤ THỂ TẾ BÀO T - VAI TRÒ TRONG ĐÁP ỨNG MIỄN DỊCH
Gene Mayer, Ph.D
Emertius Professor of Pathology, Microbiology and Immunology
University of South Carolina
Jennifer Nyland, Ph.D
Assistant Professor of Pathology, Microbiology and Immunology
University of South Carolina
Biên dịch:
Nguyễn Văn Đô, MD., PhD.,
Bộ môn Sinh lý bệnh-Miễn dịch,
Trường Đại học Y Hà Nội,
Hà Nội, Việt Nam
|
|
TURKISH |
|
FRANCAIS |
|
PORTUGUESE |
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|

 |
|
Logo image © Jeffrey
Nelson, Rush University, Chicago, Illinois and
The MicrobeLibrary |
|
|
|
|
MỤC TIÊU GIẢNG DẠY
Giới thiệu tổng quan về vai trò của các phức hợp hòa hợp
mô chủ yếu trong đáp ứng miễn dịch.
Mô tả cấu trúc và chức năng của các phân tử MHC lớp I và lớp II.
Thảo luận về bản chất của sự đa hình trong phân tử MHC lớp I và lớp II.
Mô tả cấu trúc thụ thể tế bào T đối với kháng nguyên.
Thảo luận về các cơ sở di truyền về sự đa dạng của TCR.
Thảo luận về vai trò của các phức hợp CD3 và các phân tử đồng kích thích.
Mô tả bản chất của các synap miễn dịch.
Thảo luận về các yêu cầu cho hoạt hóa tế bào T.
|
TỔNG QUAN LỊCH SỬ
Sự tương tác giữa tế bào với tế bào trong
đáp ứng miễn dịch đặc hiệu là rất quan trọng để bảo vệ cơ thể khỏi tác nhân gây
bệnh. Các tương tác này được hình thành bởi các synap miễn dịch mà các thành
phần chủ yếu là thụ thể kháng nguyên của tế bào T (TCR) và phân tử phức hợp hòa
hợp mô chủ yếu (MHC). Chức năng chính của TCR là để nhận biết kháng nguyên nằm
trong phân tử MHC và để truyền tín hiệu kích thích vào bên trong tế bào. Vì liên
kết của peptid với MHC là không đồng hóa trị, nên có nhiều yếu tố giúp ổn định
synap miễn dịch.
Sản phẩm gen mã hóa cho MHC lần đầu tiên
được phát hiện là có vai trò quan trọng trong việc loại bỏ các mô ghép. Hơn nữa,
các gen trong MHC đã được tìm thấy là rất đa hình (tức là trong quần thể đã có
nhiều loại allele khác nhau của gen). Những nghiên cứu ở chuột thuần chủng cho
thấy các gen trong MHC cũng tham gia trong việc kiểm soát cả hai đáp ứng miễn
dịch dịch thể và miễn dịch qua trung gian tế bào. Ví dụ, một số chủng chuột có
đáp ứng với một kháng nguyên đặc biệt, nhưng chủng chuột khác không thể và các
chủng này chỉ khác nhau một hoặc nhiều gen MHC. Nghiên cứu sau đó cho thấy rằng
có hai loại phân tử được mã hóa bởi các gen MHC – Các phân tử lớp I và lớp II,
chúng được nhận biết bởi các dưới lớp tế bào T khác nhau. Các phân tử lớp I được
phát hiện trên tất cả các tế bào có nhân (không có trên hồng cầu) trong khi các
phân tử lớp II được tìm thấy chỉ trên các tế bào trình diện kháng nguyên (APC),
chúng bao gồm các tế bào có tua, các đại thực bào, tế bào B và một vài loại tế
bào khác (Hình 1).
Chỉ đến khi phát hiện ra cách TCR nhận ra
kháng nguyên thì vai trò của gen MHC trong đáp ứng miễn dịch mới được hiểu rõ.
TCR có chức năng nhận biết kháng nguyên peptid trong phức hợp với các phân tử
MHC. Các tế bào T nhận ra phần kháng nguyên protein được liên kết không đồng hóa
trị với các sản phẩm gen MHC. Các tế bào T độc tế bào (Tc) nhận biết peptid liên
kết với các phân tử MHC lớp I và các tế bào T trợ giúp (Th) nhận biết peptid
liên kết với các phân tử MHC lớp II. Cấu trúc không gian ba chiều của các phân
tử MHC và TCR đã được xác định bằng X-quang tinh thể để tạo ra một bức tranh rõ
ràng về cách TCR, sản phẩm gen MHC và kháng nguyên tương tác với nhau.
|
 Hình 1
Hình 1
Phân bố của các phân tử MHC lớp I và lớp II trên tế bào người
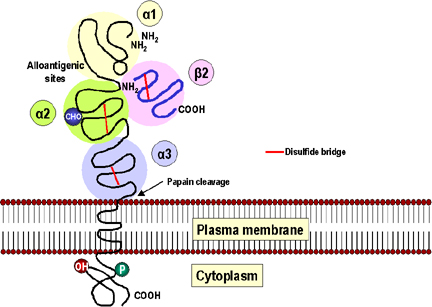
 Hình 2
Hình 2
Phân tử MHC lớp I có ba vùng hình cầu alpha 1 (màu vàng), alpha 2 (màu
xanh lá cây) và alpha 3 (màu xanh da trời). Vùng alpha 3 liên kết chặt
chẽ với các microglobulin beta 2 được mã hóa không do gen MHC (màu hồng).
Sau đó được ổn định bằng một cầu disulfid (màu đỏ) và nó cũng tương tự
như một vùng kháng thể trong cấu trúc ba chiều. Các vị trí có tính kháng
nguyên đồng loài chứa các quyết định kháng nguyên đặc hiệu cho từng cá
thể được tìm thấy ở các vùng 1 và 2. Tiếp theo có một chuỗi carbohydrat
(màu xanh, CHO). Có một phosphate nằm trong vùng bào tương. Papain sẽ
cắt gần bề mặt ngoài của màng
|
CẤU TRÚC CỦA CÁC PHÂN TỬ MHC LỚP I
Phân tử Các
phân tử MHC lớp I gồm hai chuỗi polypeptid, một chuỗi dài α và một
chuỗi ngắn β gọi là β2-microglobulin (Hình 2). Chuỗi α có bốn vùng.
- Một vùng bào tương có chứa các vị trí
phosphoryl hóa và liên kết với các thành phần khung của tế bào
- Một vùng xuyên màng có chứa các acid amin kỵ
nước mà phân tử được gắn vào màng tế bào.
- Một vùng giống kháng thể α3 được bảo tồn để
cho CD8 gắn vào.
- Một vùng gắn peptid có tính đa hình cao được
hình thành từ các vùng α1 và α2. β2-microglobulin liên kết với
chuỗi α và giúp duy trì cấu hình thích hợp của phân tử.
Các rãnh gắn kháng nguyên
Khi phân tích phân tử MHC lớp I để tìm xem
phần nào của phân tử là vùng thay đổi nhất, thì người ta thấy
rằng các vùng α1 và α2 là biến đổi nhiều nhất, nó bao gồm vùng
liên kết peptid (Hình 3). Cấu trúc của các rãnh gắn peptid, được
bộc lộ bởi X-quang tinh thể, cho thấy rãnh gồm hai vòng xoắn α
tạo thành hai bức tường ở hai bên và tám tấm gấp nếp β hình
thành một cái sàn. Peptid đựợc gắn vào rãnh và phần còn lại là
đường biên mà rãnh tạo liên kết với các peptid (Hình 4). Đây là
phần còn lại có tính đa hình nhất. Các rãnh sẽ chứa peptid dài
khoảng 8-10 acid amin. Mỗi loại peptid riêng sẽ bám vào đường
rãnh này và phụ thuộc vào các acid amin sắp xếp trong rãnh. Vì
phân tử MHC lớp I là đa hình, nên các phân tử lớp I khác nhau sẽ
liên kết với các phân tử peptid khác nhau. Mỗi phân tử lớp I chỉ
liên kết với những peptid nhất định và nó có các tiêu chuẩn mà
một peptid phải có để liên kết với các đường rãnh. Ví dụ, Hình 5
cho thấy một phân tử lớp I sẽ kết với các peptid có một leucin
(L) là acid amin nằm ở đầu tận carboxy và tyrosin (Y) hoặc
phenylalanin (F) là acid amin thứ 4 kể từ đầu tận carboxy. Khi
hai điều kiện được đáp ứng một peptid sẽ liên kết, bất kể các
acid amin khác là loại gì. Tương tự như vậy một phân tử lớp I
khác nhau sẽ gắn với bất kỳ phân tử peptid có một tyrosin (Y) là
acid amin thứ hai từ đầu tận amin và một trong hai valin (V),
isoleucin (I) hoặc leucin (L) tại đầu cuối carboxy (Hình 5). Như
vậy, cứ mỗi phân tử lớp I, có một số acid amin nhất định phải
nằm ở một vị trí đặc biệt trong peptid trước khi nó bám vào các
phân tử MHC. Những vị trí này trong peptid được gọi là "các vị
trí neo". Các đầu của peptid được che lấp trong vùng giới hạn
của rãnh gắn lớp I, trong khi đó phần trung tâm lồi ra để trình
diện cho TCR.
MHC có 6 gen mã hóa cho các phân tử lớp I là
HLA -A, HLA-B, HLA-C, HLA-E, HLA-F và HLA-G. Trong số này, HLA
A, HLA-B và HLA-C là quan trọng nhất và đa hình nhất. Bảng 1 cho
thấy mức độ đa hình tại mỗi locus này.Figur
|
 Hình 3
Hình 3
Hầu hết các biến đổi trong các acid amin tại các vị trí khác nhau trong
chuỗi alpha của phân tử MHC lớp I xảy ra trong các vùng alpha 1 và alpha
2. Sự đa hình lớn nhất được tìm thấy đối với các acid amin nằm ỏ thành
và sàn của rãnh, nơi nó liên kết với các peptid.
|
 Hình 4
Hình 4
a. Rãnh gắn peptid của phân tử MHC lớp I.
b. Rãnh có phần rất thay đổi được thể hiện rõ. Phần thay đổi được bao
bọc xung quanh bởi hốc gắn peptid.
 Hình 5
Hình 5
Các vị trí mỏ neo trong peptid liên kết với phân tử MHC lớp I (adapted
from Janeway et al. Immunobiology 6th Edition)
|
| |
|
Bảng 1. Sự đa hình của các gen MHC
lớp I |
| Locus |
Số
allele
(allotypes) |
| HLA-A |
218 |
| HLA-B |
439 |
| HLA-C |
96 |
| HLA-E, HLA-F
và HLA-G |
Các
allel ít liên quan |
|
|
|
 Hình 6
Hình 6
Các phân tử MHC lớp II gồm hai chuỗi acid amin không giống nhau (alpha
và beta), liên kết không đồng hóa trị và xuyên màng tế bào với đầu N tận
ở bên ngoài của tế bào. Các vùng gần màng tế bào nhất của mỗi chuỗi có
cấu trúc giống các kháng thể. Với ngoại lệ của vùng alpha 1, tất cả các
vùng được giữ ổn định bằng cầu disulfid (màu đỏ). Cả hai chuỗi alpha và
beta được glycosyl hóa. Các chuỗi beta ngắn hơn chuỗi alpha (trọng lượng
phân tử chuỗi beta = 28.000) và chứa các vị trí kháng nguyên dị loài.
Chuỗi alpha của một số phân tử MHC II có tính đa hình.
 Hình 7
Hình 7
Sự đa hình lớn nhất ở chuỗi beta của các phân tử MHC lớp II được tìm
thấy ở những acid amin trong vùng beta I nằm ở thành và sàn của rãnh gắn
với các peptid.
|
CẤU TRÚC CÁC PHÂN TỬ
MHC LỚP II
Phân tử
Các phân tử MHC lớp II bao gồm hai chuỗi
polypeptid, một chuỗi α và một chuỗi β có chiều dài tương đối bằng
nhau (Hình 6). Cả hai chuỗi có bốn vùng:
- Một vùng bào tương có chứa các vị trí
phosphoryl hóa và liên kết với các thành phần khung của tế bào
- Một vùng xuyên màng có chứa các acid amin kỵ
nước mà phân tử được gắn vào màng tế bào
- Có các vùng α2 và β2 bảo tồn cao cho CD4 gắn
vào
- Một vùng gắn peptid rất đa hình được hình
thành từ các vùng α1 và β1
Rãnh gắn kháng nguyên
Giống như các phân tử MHC lớp I, khi phân tích các
phần của phân tử MHC lớp II và thấy rằng vùng thay đổi nhất là α1 và
β1, trong đó có vùng liên kết peptid (Hình 7). Cấu trúc của rãnh
liên kết peptid được quan sát bởi X-quang tinh thể, và thấy rằng nó
giống như các phân tử MHC lớp I, rãnh là gồm hai chuỗi xoắn α tạo
thành bức tường ở hai bên và tám tấm gấp nếp β tạo nên một cái sàn.
Cả hai chuỗi α1 và β1 tham gia vào rãnh gắn peptid. Peptid được gắn
vào rãnh và phần còn lại giới hạn rãnh tạo liên kết với peptid. Phần
còn lại là nơi có tính đa hình nhất. Các rãnh của các phân tử lớp II
được mở ở một đầu để các rãnh có thể chứa peptid dài hơn, khoảng
13-25 axit amin với một số các acid amin nằm bên ngoài của rãnh. Một
peptid cụ thể sẽ liên kết với đường rãnh này sẽ phụ thuộc vào các
acid amin đường rãnh. Bởi vì các phân tử lớp II là đa hình, các phân
tử lớp II khác nhau sẽ liên kết peptid khác nhau. Giống như phân tử
lớp I, mỗi phân tử lớp II sẽ chỉ gắn một peptid nhất định và sẽ có
tiêu chí mà một peptid phải có để liên kết với các rãnh (nghĩa là "các
vị trí neo").
Trong MHC có 5 loci mã hóa cho các phân tử lớp II,
mỗi loci trong số đó có chứa một gen cho một chuỗi α và ít nhất một
gen cho một chuỗi β. Các locus được định rõ là HLA-DP, HLA-DQ, HLA-DR,
HLA-DM, và HLA-DO. Trong đó, HLA-DP, HLA-DQ, và HLA-DR là quan trọng
nhất và đa hình nhất. Bảng 2 cho thấy mức độ đa hình tại mỗi locus
này.
|
| |
CÁC KHÍA CẠNH QUAN TRỌNG
CỦA MHC
-
Mặc dù có tính đa hình cao của
một loài, một cá nhân có tối đa sáu lớp sản phẩm MHC lớp I khác nhau
và chỉ hơi nhiều hơn các sản phẩm MHC lớp II (chỉ xem xét các loci
chính).
-
Mỗi phân tử MHC chỉ có một vị
trí gắn. Các peptid khác nhau cho một phân tử MHC có thể gắn kết tất
cả ở cùng một vị trí, nhưng chỉ có một lần.
-
Mỗi phân tử MHC có liên kết
peptid khác nhau.
-
Tính đa hình MHC chỉ được xác
định trong dòng mầm. Không có cơ chế tái tổ hợp để tạo ra sự đa
dạng.
-
Các phân tử MHC được gắn lên
màng tế bào; nhận biết bởi các tế bào T và cần phải có tiếp xúc trực
tiếp tế bào-tế bào.
-
Các allele của các gen MHC là
đồng trội. Mỗi sản phẩm gen MHC được biểu lộ trên bề mặt tế bào có
nhân.
-
Một peptid phải liên kết với MHC
nhất định của một cá thể, nếu không sẽ không có đáp ứng miễn dịch.
Đó là mức độ kiểm soát thứ nhất.
-
Các tế bào T trưởng thành phải
có một thụ thể tế bào T để nhận ra peptid liên kết với MHC. Đây là
cấp độ kiểm soát thứ hai.
-
Các cytokin (đặc biệt là
interferon-γ) sẽ làm tăng cường mức độ biểu lộ MHC.
-
Các peptid từ bào tương liên kết
với MHC lớp I và được nhận biết bởi các tế bào Tc. Peptid liên kết
với MHC lớp II và được nhận biết bởi các tế bào Th.
-
Tính đa hình của MHC là quan
trọng cho sự sống còn của loài.
|
Bảng 2. Tính đa hình của gen MHC
lớp II |
| Locus |
Số
allele
(allotypes) |
HLA-DPA
HLA-DPB |
12
88 |
HLA-DQA
HLA-DQB |
17
42 |
HLA-DRA
HLA-DRB1
HLA-DRB3
HLA-DRB4
HLA-DRB5 |
2
269
30
7
12 |
| HLA-DM và
HLA-DO |
Allele ít
liên quan |
|
| |
LÀM THẾ NÀO ĐỂ GẮN PEPTID VÀO RÃNH MHC?
Peptid từ bào tương liên kết với MHC lớp I và được nhận
biết bởi các tế bào CTL. Các peptid đi vào mạng lưới nội chất và gắn vào
rãnh MHC lớp I. Phức hợp này sau đó được chuyển sang bề mặt tế bào thông
qua bộ Golgi. Các phân tử MHC lớp II được hình thành với một chuỗi bất
biến là (Ii) như là một người giữ chỗ ở các ER và Golgi. Chuỗi Ii bị cắt
và lấy ra một lần phức hợp ở trong túi. Peptid từ trong túi liên kết với
MHC lớp II và sau đó được chuyển ra bề mặt tế bào, nơi chúng được nhận
biết bởi các tế bào Th.
VAI TRÒ CỦA TCR TRONG ĐÁP ỨNG MIỄN DỊCH
TCR là một phân tử được tìm thấy trên bề mặt tế bào T
để nhận biết kháng nguyên được gắn trong MHC. TCR này có cấu trúc tương
tự như kháng thể và là một phần của siêu gia đình kháng thể. Có hai loại
TCR, chủ yếu là αβ và thường thấy trong các mô lympho, và γδ được phát
hiện ở bề mặt niêm mạc.
|
 Hình 8
Hình 8
Dị dimer của thụ thể tế bào T bao gồm hai glycoprotein xuyên màng, các
chuỗi alpha và beta. Có hai vùng ở phần bên ngoài của mỗi chuỗi và chúng
hình thành các vùng thay đổi và hằng định của kháng thể. Có các phân tử
đường trên mỗi vùng. Có một chuỗi ngắn tương tự như vùng bản lề của
kháng thể để nối các vùng giống kháng thể giống như chuỗi xuyên màng. Nó
có chứa cystein và hình thành một cầu nối disulfid. Các cấu trúc màng kỵ
nước xoắn ốc là không bình thường ở chỗ chúng chứa các acid amin điện
tích dương (acid amin cơ bản). Các chuỗi alpha có hai vị trí điện tích
dương, trong khi chuỗi beta có một.
 Cấu trúc của thụ thể tế bào T-A6 liên kết với phân tử MHC lớp I tạo phức
hợp với một peptid Y8a HTLV-1 Tax đã bị thay đổi. Peptid HIV được thể
hiện ở màu xám. Phân tử MHC lớp I biểu hiện ở màu xanh đậm, liên kết với
beta 2 microglobulin bởi màu xanh nhạt. Thụ thể tế bào T có màu xanh lá
cây và màu vàng.
Cấu trúc của thụ thể tế bào T-A6 liên kết với phân tử MHC lớp I tạo phức
hợp với một peptid Y8a HTLV-1 Tax đã bị thay đổi. Peptid HIV được thể
hiện ở màu xám. Phân tử MHC lớp I biểu hiện ở màu xanh đậm, liên kết với
beta 2 microglobulin bởi màu xanh nhạt. Thụ thể tế bào T có màu xanh lá
cây và màu vàng.
Y. H.Ding, B. M.Baker, D. N.Garboczi, W. E.Biddison & D. C.Wiley MMDB
Id: 11766 PDB Id: 1QSF Image prepared using RasMol
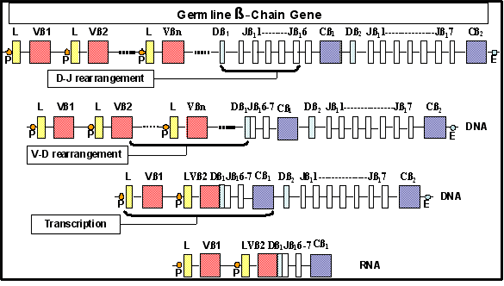
 Hình 9
Hình 9
Tái tổ hợp các gen tổng hợp chuỗi beta của TCR
|
CẤU TRÚC CỦA THỤ THỂ TẾ BÀO T (TCR)
TCR là một dị dimer gồm một chuỗi α và
một chuỗi β có chiều dài xấp xỉ bằng nhau (Hình 8). Mỗi chuỗi có một cái
đuôi ngắn nằm trong bào tương nhưng nó rất nhỏ để có thể truyền một tín
hiệu hoạt hóa cho tế bào. Cả hai chuỗi có một vùng xuyên màng bao gồm
các acid amin kỵ nước, nhờ nó mà phân tử này được neo ở màng tế bào. Cả
hai chuỗi có một vùng hằng định và một vùng thay đổi giống như các kháng
thể. Vùng thay đổi của cả hai chuỗi chứa các vùng siêu biến đổi để xác
định sự đặc hiệu kháng nguyên. Mỗi tế bào T có một TCR có tính đặc hiệu
duy nhất (ngoại trừ allele).
CƠ SỞ DI TRUYỀN CỦA SỰ HÌNH THÀNH THỤ THỂ
Cơ sở di truyền của việc hình thành các
thụ thể với kháng nguyên trên tế bào B đã được thảo luận trước đó (xem
phần về di truyền học Ig). Việc tạo ra một loạt các TCRs được thực hiện
bằng một cơ chế tương tự. Các gen tổng hợp ra β TCR được cấu thành từ V,
D và các mảnh của gen J tái sắp xếp trong quá trình phát triển tế bào T
để sản xuất chuỗi β TCR khác nhau (Hình 9). Các gen dòng mầm cho gen α
TCR được cấu thành từ V và các mảnh gen J tái sắp xếp để tạo ra các
chuỗi α. Sự đặc hiệu của TCR được xác định bởi sự kết hợp của các chuỗi
α và β.
Có một quần thể nhỏ các tế bào T biểu lộ
TCR có chuỗi γ và δ thay vì chuỗi α và β. Những tế bào T gamma và delta
chiếm ưu thế trong các biểu mô niêm mạc và có khả năng chống kháng
nguyên vi khuẩn và virút. Các gen cho các chuỗi δ có các gen V, D và J
trong khi các gen cho các chuỗi γ chỉ có gen V và J nhưng đội quân này
là nhỏ hơn đáng kể so với các tế bào T alpha/beta. Các tế bào T
gamma/delta nhận biết kháng nguyên trong MHC một cách độc lập không
giống như tế bào T alpha/beta.
CÁC KHÍA CẠNH QUAN TRỌNG CỦA
TCR
-
Mỗi tế bào T chỉ mang một TCR đặc
hiệu (có loại trừ allele).
-
Các TCR αβ chỉ nhận biết kháng nguyên
trong điều kiện có tương tác tế bào với tế bào và kháng nguyên nằm
trong MHC.
-
Các TCR γδ nhận biết kiểu kháng
nguyên không phụ thuộc MHC để đáp ứng với kháng nguyên của virút và
vi khuẩn nhất định.
|
Bảng 3
SO SÁNH CÁC ĐẶC ĐIỂM CHÍNH CỦA CÁC GEN KHÁNG THỂ VÀ THỤ THỂ TẾ BÀO T VÀ
CÁC PROTEIN
|
|
GEN |
|
Đặc điểm |
Ig |
TCR |
|
Nhiều VDJs, một vài C's |
Có |
Có |
|
Tái sắp xếp VDJ |
Có |
Có |
|
Cặp V hình thành vị trí nhận biết KN |
Có |
Có |
|
Siêu đột biến soma |
Có |
Không |
|
PROTEIN |
|
Dạng xuyên màng |
Có |
Có |
|
Dạng tiết |
Có |
Không |
|
Isotype với chức năng riêng biệt |
Có |
Không |
|
Hóa trị |
2 |
1 |
|
Trích từ Miễn dịch học Janeway
and Travers |
|
 Cấu trúc dạng tinh thể của một phức hợp gồm một thụ thể
tế bào T của người, peptid kháng nguyên cúm Ha và một phân tử MHC lớp
II. Các chuỗi alpha và beta của các phân tử MHC II được thể hiện bởi màu
đen và màu xanh nhạt. Các thụ thể tế bào T có màu vàng và xanh lá cây.
Các peptid cúm là màu xám. Hennecke, J., Carfi, A., Wiley, D. C. MMDB
Id: 14648 PDB Id: 1FYT. Image prepared using RasMol
Cấu trúc dạng tinh thể của một phức hợp gồm một thụ thể
tế bào T của người, peptid kháng nguyên cúm Ha và một phân tử MHC lớp
II. Các chuỗi alpha và beta của các phân tử MHC II được thể hiện bởi màu
đen và màu xanh nhạt. Các thụ thể tế bào T có màu vàng và xanh lá cây.
Các peptid cúm là màu xám. Hennecke, J., Carfi, A., Wiley, D. C. MMDB
Id: 14648 PDB Id: 1FYT. Image prepared using RasMol |
|
|
 Hình 10
Hình 10
Thụ thể kháng nguyên trên bề mặt tế bào T bao gồm tám protein.
(a) Hai chuỗi liên kết với nhau bằng cầu nối disulfua của thụ thể tế bào
T hình thành nên một dị dimer. Chúng nhận biết các peptid đã được đặt
trong các phân tử MHC.
(b) Bốn chuỗi, gọi chung là CD3, liên kết với dimer thụ thể tế bào T và
tham gia vận chuyển của nó tới bề mặt của tế bào. Phức hợp CD3 cùng với
các chuỗi zeta, hình thành nên một đồng dimer, truyền tín hiệu sau khi
kháng nguyên gắn kết.
|
TCR VÀ PHỨC HỢP CD3
TCR có liên quan gần gũi với một
nhóm 5 protein được gọi chung là phức hợp CD3 (Hình 10). Các phức hợp
CD3 bao gồm một chuỗi γ, một chuỗi δ, hai chuỗi ε và 2 chuỗi ξ. Tất cả
các protein của phức hợp CD3 là bất biến và chúng không đóng góp vào độ
đặc hiệu. Các phức hợp CD3 là cần thiết để biểu lộ TCR trên bề mặt tế
bào trong quá trình phát triển của tế bào T. Ngoài ra, phức hợp CD3
truyền tín hiệu hoạt hóa vào trong tế bào sau khi KN tương tác với TCR.
|
 Hình 11
Hình 11
A. Các phân tử liên quan đến sự tương tác giữa các tế bào T và tế bào
trình diện kháng nguyên. Một số cytokin được sản xuất bởi mỗi loại tế
bào đã được thể hiện.

B. Phối tử có liên quan đến sự tương tác của
các tế bào T độc tế bào và các tế bào đích của chúng
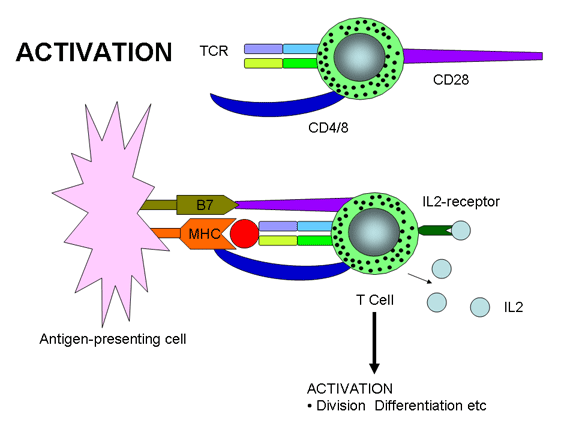
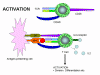 Hình 12a
Hình 12a
Hoạt hóa tế bào T chỉ xảy ra khi cả hai TCR và các phân tử đồng kích
thích được kết hợp với các phối tử tương ứng của chúng
 Hình 12b
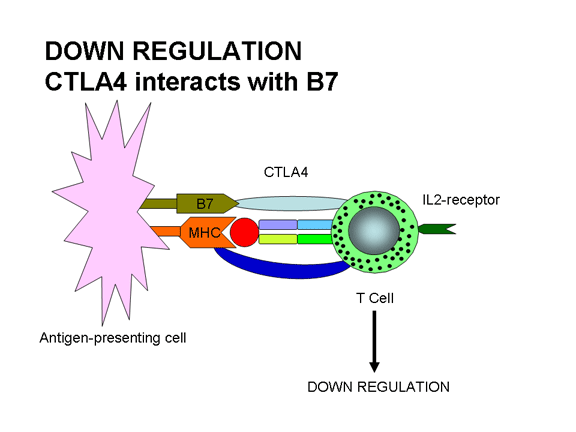
Hình 12b
Sự giảm điều hòa xuất hiện nếu CTLA-4 tương tác với B7: CTLA-4 gửi một
tín hiệu ức chế
 Hình 12c
Hình 12c
Phối hợp của TCR và kháng nguyên / MHC trong trường hợp không có mặt của
đồng kích thích có thể dẫn đến suy giảm
 Hình 12d
Hình 12d
Tham gia của các phân tử đồng kích thích trong trường hợp không có TCR
dẫn đến không có đáp ứng
|
SYNAP MIỄN DỊCH
Sự tương tác giữa các phân tử TCR và MHC là không
mạnh mẽ. Các phân tử phụ là rất cần thiết để giúp ổn định các tương tác (Hình
11a, b). Chúng bao gồm:
-
CD4 liên kết với MCH lớp II, nó đảm bảo rằng
các tế bào Th chỉ tương tác với APC
-
CD8 liên kết với MHC lớp I, đảm bảo rằng các
tế bào Tc có thể tương tác với các tế bào đích
-
CD2 liên kết với LFA-3
-
LFA-1 liên kết với ICAM-1
Các phân tử phụ là bất biến và không đóng góp vào
sự đặc hiệu của sự tương tác, được nhận biết bởi TCR. Sự biểu lộ các phân tử phụ
có thể được tăng lên khi có cytokin, là một cách mà các cytokin có thể điều hòa
đáp ứng miễn dịch.
Ngoài các phân tử phụ giúp ổn định sự tương tác giữa các TCR và kháng nguyên gắn
với các phân tử MHC, các phân tử khác cũng cần thiết để hoạt hóa tế bào T. Hai
tín hiệu cần thiết để hoạt hóa tế bào T - một là sự kết hợp của TCR với Ag/MHC
và hai là tín hiệu khác từ sự tham gia của các phân tử đồng kích thích với các
phối tử của chúng. Một trong những phân tử đồng kích thích quan trọng nhất (nhưng
không chỉ có mình nó) là CD28 trên tế bào T, nó phải tương tác với B7-1 (CD80)
hoặc B7-2 (CD81) trên các APC. Giống như các phân tử phụ các phân tử đồng kích
thích là bất biến và không đóng góp vào sự đặc hiệu của sự tương tác. Các đa
tương tác của TCR với Ag /MHC và các phân tử phụ và đồng kích thích với các phối
tử của họ được gọi là "miễn dịch synap”.
Không chỉ có sự đồng kích thích là cần thiết để
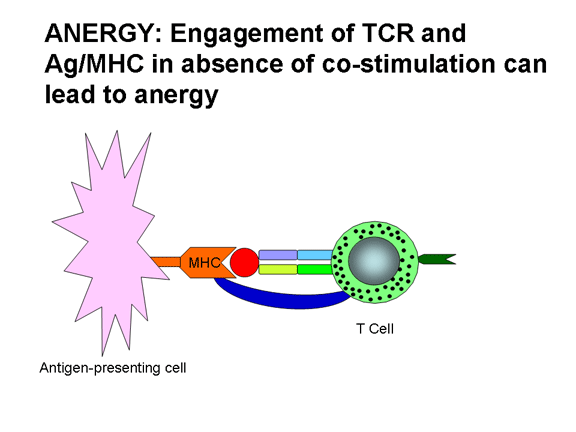
hoạt hóa tế bào T, thiếu một kích thích phù hợp có thể dẫn đến suy giảm (tức là,
mất khả năng đáp ứng với kháng nguyên) hoặc đáp ứng giảm xuống. Hình 12 cho thấy
các kết quả có thể có của một tế bào T nhận được một hoặc cả hai tín hiệu cần
thiết để hoạt hóa. Có sự kết hợp của TCR với Ag/MHC nhưng không có đồng kích
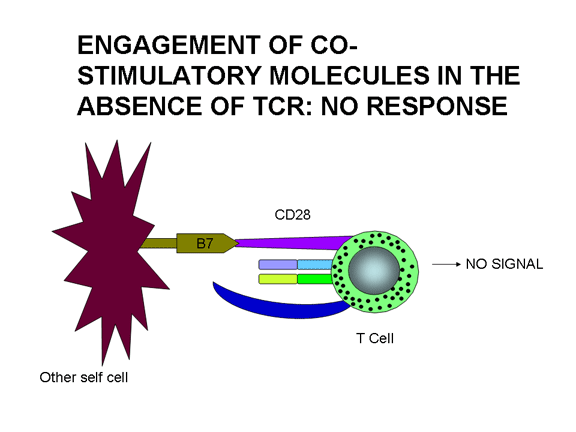
thích dẫn đến suy giảm. Nếu chỉ có các phân tử đồng kích thích tham gia thì
không có tác dụng. Sự kết hợp của TCR với Ag/MHC và các phân tử đồng kích thích
với các kết quả phối tử dẫn đến sự hoạt hóa. Sự liên kết của TCR với Ag/MHC và
sự tham gia của phối tử B7 với CTLA-4, một phân tử tương tự như CD28, kết quả là
giảm đáp ứng. Tương tác CTLA-4/B7 sẽ gửi một tín hiệu ức chế cho các tế bào T
chứ không phải là một tín hiệu kích thích. Đây là một trong những cách để điều
hòa đáp ứng miễn dịch. CTLA-4 được biểu lộ muộn hơn trên các tế bào T trong một
đáp ứng miễn dịch và điều này sẽ giúp cho việc dừng đáp ứng.
CÁC BƯỚC CHÍNH CỦA HOẠT HÓA TẾ BÀO T
-
APC phải xử lý và trình diện peptid cho các
tế bào T
-
Các tế bào T phải nhận được một tín hiệu đồng
kích thích - thường là từ CD28/B7
-
Các phân tử bám dính phụ phải giúp đỡ để ổn
định cho sự liên kết của tế bào T và APC. (CD4/MHC lớp II, CD8/ MHC lớp I,
LFA-1/ICAM-1 và CD2/LFA-3)
-
Tín hiệu từ bề mặt tế bào phải được chuyển
đến nhân thông qua tín hiệu thứ hai
-
Cytokin, bao gồm IL-2, phải giúp tế bào phân
chia
|
Bảng 4
Các phân tử phụ quan trọng |
|
Phân tử ở tế bào T
|
Phân tử ở tế bào thứ 2 |
|
CD4 trên tế bào T trợ giúp |
Các phân tử MHC lớp I |
|
CD8 trên tế bào T độc |
Các phân tử MHC lớp II |
|
LFA-2 (CD2) |
LFA-3 |
|
LFA-1 |
ICAM-1, ICAM-2 |
|
LFA = Kháng nguyên |
|
ICAM = Phân tử kết dính gian bào |
|
|
|
 Trở về phần Miễn dịch của Vi khuẩn học và Miễn dịch học
online
Trở về phần Miễn dịch của Vi khuẩn học và Miễn dịch học
online
This page last changed on
Thursday, September 14, 2017
Page maintained by
Richard Hunt
|
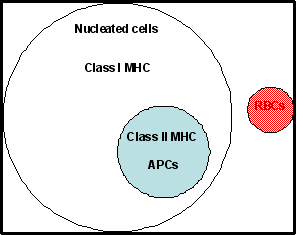 Hình 1
Hình 1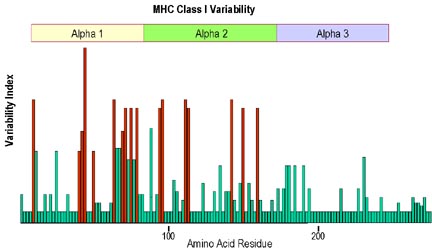 Hình 3
Hình 3 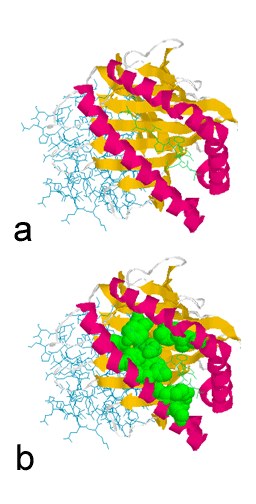 Hình 4
Hình 4 Hình 6
Hình 6 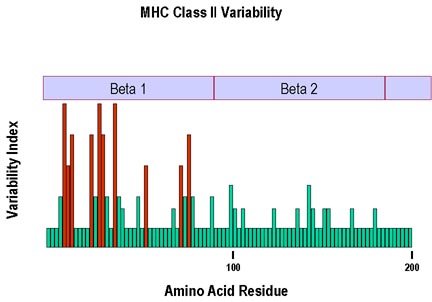 Hình 7
Hình 7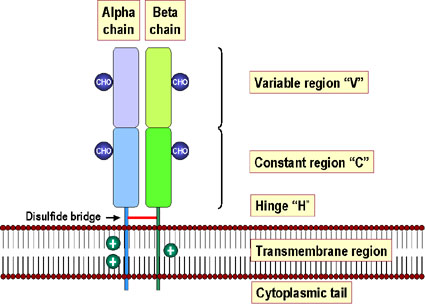 Hình 8
Hình 8 Cấu trúc dạng tinh thể của một phức hợp gồm một thụ thể
tế bào T của người, peptid kháng nguyên cúm Ha và một phân tử MHC lớp
II. Các chuỗi alpha và beta của các phân tử MHC II được thể hiện bởi màu
đen và màu xanh nhạt. Các thụ thể tế bào T có màu vàng và xanh lá cây.
Các peptid cúm là màu xám. Hennecke, J., Carfi, A., Wiley, D. C. MMDB
Id: 14648 PDB Id: 1FYT. Image prepared using RasMol
Cấu trúc dạng tinh thể của một phức hợp gồm một thụ thể
tế bào T của người, peptid kháng nguyên cúm Ha và một phân tử MHC lớp
II. Các chuỗi alpha và beta của các phân tử MHC II được thể hiện bởi màu
đen và màu xanh nhạt. Các thụ thể tế bào T có màu vàng và xanh lá cây.
Các peptid cúm là màu xám. Hennecke, J., Carfi, A., Wiley, D. C. MMDB
Id: 14648 PDB Id: 1FYT. Image prepared using RasMol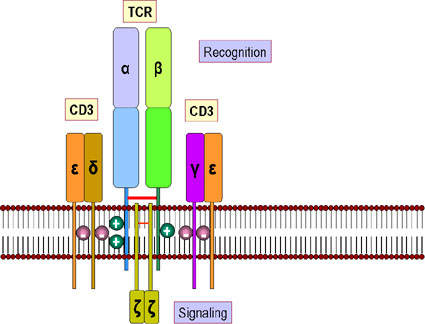 Hình 10
Hình 10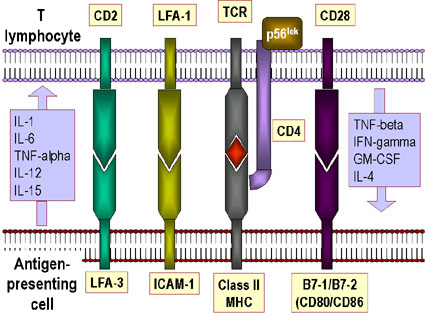 Hình 11
Hình 11