|
TEACHING OBJECTIVES
To highlight the major cytokines
that are mediators of: (i) natural immunity, (ii) adaptive immunity and
(iii) hematopoesis.
To discuss regulation of immune responses.
KEY WORDS
Monokines, Lymphokines, Interleukins, Chemokines, Redundancy, TNF-α,
IL-1, IL-10, IL-12, Interferons, IFN-γ, IL-2, IL-4, IL-5, TGF-β, GM-CSF,
M-CSF, G-CSF, Tregs
 Figure
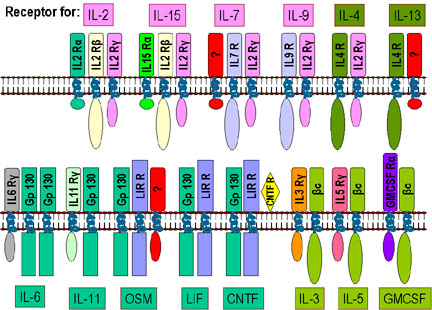
1A Receptors for various cytokines showing common subunits Figure
1A Receptors for various cytokines showing common subunits
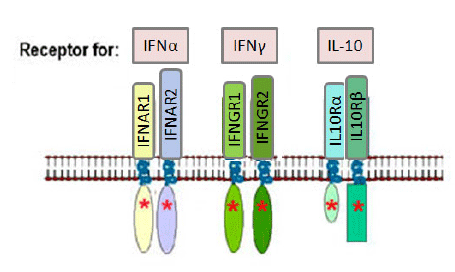
 Figure 1B Interferon receptor family
Figure 1B Interferon receptor family |
Overview
Cytokines
are a diverse group of non-antibody proteins that act as mediators between
cells. They were initially identified as products of immune cells that act as
mediators and regulators of immune processes but many cytokines are now known to
be produced by cells other than immune cells and they can have effects on
non-immune cells as well. Cytokines are currently being used clinically as
biological response modifiers for the treatment of various disorders. The term
cytokine is a general term used to describe a large group of proteins but there
are other terms that are commonly used to describe particular kinds of
cytokines. These include:
-
Monokines, cytokines produced by mononuclear
phagocytic cells
-
Lymphokines, cytokines produced by activated lymphocytes,
especially Th cells
-
Interleukins, cytokines that act as mediators
between leukocytes
-
Chemokines, small cytokines primarily
responsible for leucocyte migration
Cytokines function as part of a larger
inter-related system of proteins and signaling cascades, the cytokine network.
These are complex interactions in which different cells can respond differently
to the same cytokine depending upon other signals received by the cell. Cytokine
signaling is very flexible and can induce both protective and damaging
responses. One cytokine often influences the synthesis of other cytokines. They
can produce cascades, or enhance or suppress production of other cytokines. In
addition, they can often influence the action of other cytokines. The effects
can be: antagonistic, additive, or synergistic.
Cytokines are not typically stored as preformed proteins. Rather their
synthesis is initiated by gene transcription and their mRNAs are short lived.
They are produced as needed in immune responses. Genes encoding cytokines
can produce variants through alternative splicing to yield proteins with
slightly different but biologically significant bioactivities.
Many individual cytokines are produced by many
cell types involved in both the innate and adaptive immune response. Individual
cytokines also act on many cell types (i.e., they are
pleotropic) and in many cases cytokines have similar actions (i.e.,
they are redundant). Redundancy is due to the nature of the cytokine
receptors.
Receptors for cytokines are heterodimers (sometimes
heterotrimers) that can be grouped into families based on common structural
features; one subunit is common
to all members of a given family. Some examples are shown in Figure 1.
-
Type 1 cytokine receptors (IL-2R family) are
the largest family of cytokine receptors. This family is divided into three
subsets based on common components: IL2Rγ, common β, and gp130 (Figure 1A).
These receptors lack intrinsic protein tyrosine kinase activity. Ligand
(cytokine) binding leads to receptor dimerization and initiation of
intracellular signaling.
-
Type 2 cytokine receptors (IFNR family) have
conserved cysteines in the extracellular domains of the subunits. The
extracellular domains also have tandem immunoglobulin-like domains
characteristic of this cytokine receptor family. These receptor subunits
also have intrinsic tyrosine kinase activity (denoted by the * in Figure
1B).
Chemokine receptors all have seven transmembrane
segments linked to GTP-binding proteins. They are selectively expressed on
particular lymphocyte populations and are named based on the family of
chemokines to which they bind; CCR (the CC receptor) binds CC chemokines as its
ligand while the CXCR binds CXC chemokines as its ligand (chemokines naming
convention will be discussed below).
Since
the subunit common to all members of the family functions in binding cytokine
and in signal transduction, a receptor for one cytokine can often respond to
another cytokine in the same family. Thus, an individual lacking IL-2, for
example, is not adversely affected because other cytokines (IL-15, IL-7, IL-9,
etc.) assume its function. Similarly, a mutation in a cytokine receptor subunit
other than the one in common often has little effect. On the other hand, a
mutation in the common subunit has profound effects. For example, a mutation in
the gene for the IL-2R gamma subunit causes human X-linked severe combined
immunodeficiency (XSCID) characterized by a complete or nearly complete T and B
cell defects.
Cytokines bind to specific receptors on target cells with high affinity and
the cells that respond to a cytokine are either:
-
The same cell that secreted
cytokine (autocrine)
-
A nearby cell (paracrine)
-
A distant cell reached
through the circulation (endocrine). Cellular responses to cytokines are
generally slow (hours) because they require new mRNA and protein synthesis.
|
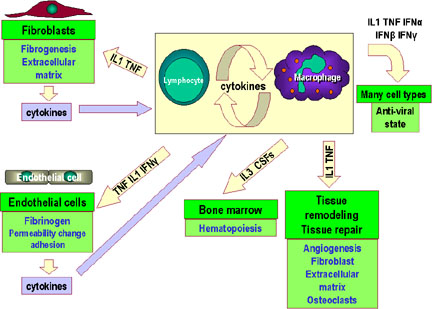
 Figure 2
Figure 2
Immunoregulatory actions of interferon gamma on the immune
system. Note the anti-proliferation and anti-ral activities are
weaker than those of IFN alpha and IFN beta. IFN gamma is the most
potent of the three at macrophage activation and in inducing class
II MHC expression |
Categories of Cytokines
Cytokines can be grouped into different categories based on their functions
or their source but it is important to remember that because they can be
produced by many different cells and act on many different cells, any
attempt to categorize them will be subject to limitations.
Mediators of natural immunity
(innate immune response)
Cytokines that play a major role in the innate immune system include:
TNF-α, IL-1, IL-10, IL-12, type I interferons (IFN-α and IFN-β), IFN-γ,
and chemokines.
TNF-α
Tumor necrosis factor alpha is produced by activated macrophages is
response to microbes, especially the lipopolysaccharide (LPS) of
Gram negative bacteria. It is an important mediator of acute
inflammation. It mediates the recruitment of neutrophils and
macrophages to sites of infection by stimulating endothelial cells
to produce adhesion molecules and by producing chemokines which are
chemotactic cytokines. TNF- α also acts on the hypothalamus to
produce fever and it promotes the production of acute phase
proteins.
IL-1
Interleukin 1 is another inflammatory cytokine produced by activated
macrophages. Its effects are similar to that of TNF-α and it also
helps to activate T cells.
IL-10
Interleukin 10 is produced by activated macrophages and Th2 cells.
It is predominantly an inhibitory cytokine. It inhibits production
of IFN-γ by Th1 cells, which shifts immune responses toward a Th2
type. It also inhibits cytokine production by activated macrophages
and the expression of class II MHC and co-stimulatory molecules on
macrophages, resulting in a dampening of immune responses.
IL-12
Interleukin 12 is produced by activated macrophages and dendritic
cells. It stimulates the production of IFN-γ and induces the
differentiation of Th cells to become Th1 cells. In addition, it
enhances the cytolytic functions of Tc and NK cells.
Type I interferons
Type I interferons (IFN-α and IFN-β) are produced by many cell types
and they function to inhibit viral replication in cells. They also
increase expression of class I MHC molecules on cells making them
more susceptible to killing by CTLs. Type I interferons also
activate NK cells.
INF-γ
Interferon gamma is an important cytokine produced by primarily by
Th1 cells, although it can also be produced by Tc and NK cells to a
lesser extent. It has numerous functions in both the innate and
adaptive immune systems as depicted in Figure 2.
Chemokines
Chemokines are chemotactic cytokines produced by many kinds of
leukocytes and other cell types. They represent a large family of
molecules that function to recruit leukocytes to sites of infection
and play a role in lymphocyte trafficking by determining which cells
will cross the epithelium and where they are directed to go. There
are four families of chemokines based on spacing of conserved
cysteine. Two examples are the α-chemokines which have a CXC
structure (two cysteines with a different amino acid in between) and
the β-chemokines which have a CC structure (two neighboring
cysteines). Individual chemokines (within the same family) often
bind more than one receptor.
|
 Figure 3
Figure 3
Immuno-regulatory actions of interleukin-2
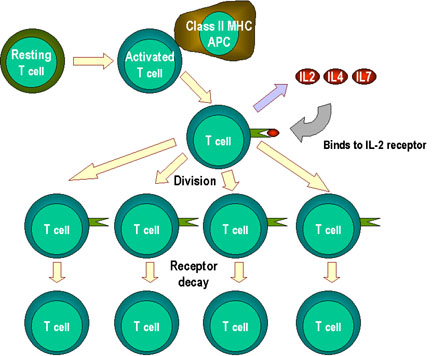
 Figure 4
Figure 4
T cell proliferation and cytokines. When T cells are
resting, they do not make cytokines such as interleukins 2, 4 or 7. Nor
do they express large amounts of their receptors. There are no IL-2
receptors. Activation of T cells results in the formation of high
affinity IL-2 receptors and induction of the synthesis and secretion of
IL-2 and Il-4. These bind to their receptors and the cells proliferate.
When stimulation by interleukins declines (e.g. when antigen stimulation
declines), receptors decay and the proliferative phase is at an
end. Note: stimulation by the cytokines can be
paracrine or
autocrine
|
Mediators of adaptive immunity
Cytokines that play a major role in the adaptive immune system include:
IL-2, IL-4, IL-5, TGF-β, IL-10 and IFN-γ.
IL-2
Interleukin 2 is produced by Th cells, although it can also be produced
by Tc cells to a lesser extent. It is the major growth factor for T
cells. It also promotes the growth of B cells and can activate NK cells
and monocytes as depicted in Figure 3. IL-2 acts on T cells in an
autocrine fashion. Activation of T cells results in expression of IL-2R
and the production of IL-2. The IL-2 binds to the IL-R and promotes cell
division. When the T cells are no longer being stimulated by antigen,
the IL-2R will eventually decay and the proliferative phase ends Figure
4.
IL-4
Interleukin 4 is produced by macrophages and Th2 cells. It stimulates
the development of Th2 cells from naïve Th cells and it promotes the
growth of differentiated Th2 cells resulting in the production of an
antibody response. It also stimulates Ig class switching to the IgE
isotype.
IL-5
Interleukin 5 is produced by Th2 cells and it functions to promote the
growth and differentiation of B cells and eosinophiles. It also
activates mature eosinophiles.
TGF-β
Transforming growth factor beta is produced by T cells and many other
cell types. It is primarily an inhibitory cytokine. It inhibits the
proliferation of T cells and the activation of macrophages. It also acts
on PMNs and endothelial cells to block the effects of pro-inflammatory
cytokines.
Stimulators of hematopoesis
Some cytokines stimulate the differentiation of hematopoetic cells. These
include GM-CSF which promotes the differentiation of bone marrow
progenitors, M-CSF, which promotes growth and differentiation of progenitors
into monocytes and macrophages and G-CSF, which promotes production of PMNs.
Interleukin 17
IL-17 is proinflammatory cytokine approximately 150 amino acids long. The
IL-17 family includes six members which share sequence homology but
differential tissue expression. IL-17 is produced by Th17 cells and its over
expression has been associated with autoimmune disease including multiple
sclerosis, rheumatoid arthritis, and inflammatory bowel disease.
|
|
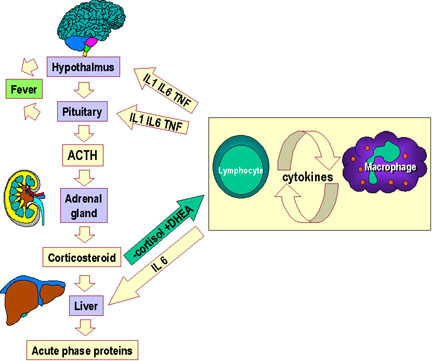
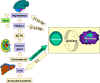 Figure 5a Cytokine network. Communication between lymphocytes and macrophages
and the hypothalamus, adrenals and the liver
Figure 5a Cytokine network. Communication between lymphocytes and macrophages
and the hypothalamus, adrenals and the liver
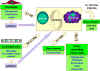 Figure 5b
Figure 5b
Cytokine network. Communication between lymphocytes and macrophages
and other cells and tissues |
Cytokine Networks
Although the focus of most research has been on the production and
action of cytokines on cells of the immune system, it is important to
remember that many of them have effects on other cells and organ
systems.In fact, the cytokine network is rather complex and represents a
series of overlapping and inter-related connections amongst cytokines.
Within this network, one cytokine may induce or suppress its own
synthesis, induce or suppress the synthesis of other cytokines, induce
or suppress synthesis of cytokine receptors (both its own and other
cytokine receptors), and antagonize or synergize with other cytokines. A diagram
showing some of the interactions in the cytokine network is presented in
Figure 5a, b and c.
|
|
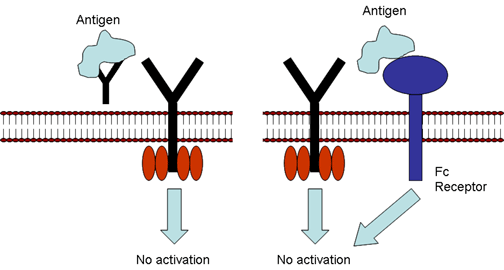
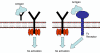 Figure 6
Figure 6
Regulation by antibody. Soluble antibody competes with cell surface Ig
for binding to antigen (left) or soluble antibody binds to Fc receptor
resulting in an inhibitory signal (right). |
Immunoregulation
The magnitude of an immune response is determined by the balance between
antigen-driven activation of lymphocytes and negative regulatory
influences that prevent or dampen the response. Regulatory mechanisms
can act at the recognition, activation or effector phases of an immune
response. Examples of regulation that have already been discussed
include:
- Recognition of antigen in the absence of co-stimulation resulting in
anergy
- Recognition of antigen with CTLA-4 engagement of B7 resulting in down
regulation of T cell activation
- Cytokines with stimulatory or inhibitory activities on immune cells
- Idiotype/anti-idiotype interactions leading to stimulation or inhibition
of immune responses
- Dose and route of antigen exposure can induce differential Th responses
which in one case can protect and in another can tolerize.
In addition to these there are other ways in which immune responses can
be regulated.
Regulation by antibody (Figure
6)
Soluble antibody can compete with antigen receptors on B cells and block
or prevent B cell activation. In addition antigen antibody complexes can
bind to Fc receptors on B cells, sending an inhibitory signal to B
cells. In this case the regulation is occurring at the recognition
level.
In addition, antigen-antibody complexes can bind to Fc receptors on B
cells, sending an inhibitory signal to B cells. Here regulation occurs
at the activation level.
Antibody can also regulate activation (enhance) by maintaining a
source of antigen for APC. In this case, antibody binds antigen forming
an immune complex which binds and activated the complement system.
Complement activation allows for ligation to the complement receptor on
the APC.
Regulation by cytokines
Cytokines are positive or negative regulators. They act at many stages
of the immune response, but their activity is dependent upon the other
cytokines present in the microenvironment as well as receptor expression
on effector cells. Cytokines regulate the type and extent of the immune
response generated.
Regulation by regulatory T cells (Tregs)
Regulatory T cells (Tregs) are a recently described populations of cells
that can regulate immune responses. They do not prevent initial T cell
activation; rather, they inhibit a sustained response and prevent
chronic and potentially damaging responses. They do not have
characteristics of Th1, Th2 or TH17 cells but they can suppress both Th1 and
Th2 responses.
Naturally occurring Tregs
The thymus gives rise to
CD4+/CD25+/Foxp3+ cells that functions as Tregs. These Tregs
suppress immune responses in a cell contact dependent manner but the
mechanism of suppression has not been established.
Induced Tregs
In the periphery some T cells are induced to
become Tregs by antigen and either IL-10 or TGF-β. Tregs induced by
IL-10 are CD4+/CD25+/Foxp3- and are referred to as Tr1 cells. These
cells suppress immune responses by secretion of IL10. Tregs induced
by TGF-β are CD4+/CD25+/Foxp3+ and are referred to as induced Tregs.
These cells suppress by secretion of TGF-β
CD8+ Tregs
Some CD8+ cells can also be induced by antigen and
IL-10 to become a Treg cell. These cells are CD8+/Foxp3+ and they
suppress by a cell contact dependent mechanism or by secretion of
cytokines. These cells have been demonstrated in vitro but it is not
known whether they exist in vivo.
Genetic factors influencing
immunoregulation
MHC-linked genes help control response to infection. Certain HLA
haplotypes are associated with individuals who are responders or
nonresponder, those who are susceptible or resistant.
Non-MHC genes are also involved in immunoregulation. An example is a
gene related to macrophage activity encoding a transporter protein
involved in transport of nitrite (NO2-) into the phagolysosome, natural
resistance-associated macrophage protein-1 (Nramp1). Polymorphisms in
this gene could change the activity of macrophages.
Cytokine, chemokine, and their receptors are involved in
immunoregulation as discussed above. Polymorphisms in the genes encoding
these, in particular the receptors, have been shown to correlate to
susceptibility to infection or generation of autoimmune disease.
|

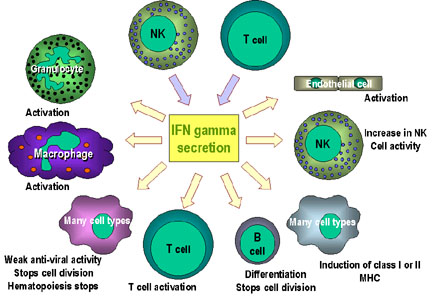 Figure 2
Figure 2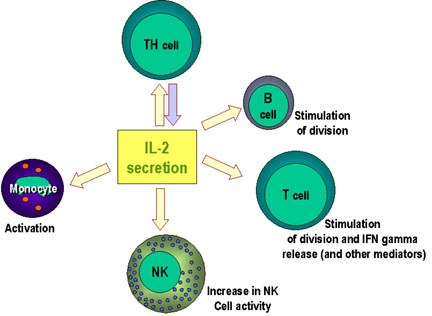 Figure 3
Figure 3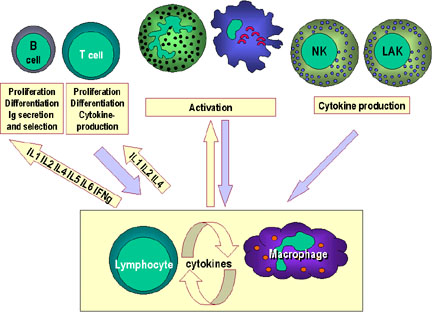 Figure 5c
Figure 5c