![]()

BACTERIOLOGY - CHAPTER ONE
THE BACTERIAL CELL
Dr Alvin Fox
Emeritus Professor
University of South Carolina School of Medicine
Let us know what you think
FEEDBACK
![]()
Logo image © Jeffrey Nelson, Rush University, Chicago, Illinois and The MicrobeLibrary
KEY WORDS
Prokaryotic
Eubacteria (Bacteria)
Archaebacteria (Archaea)
Eukaryotic
Plasmid
Chromosome
Ribosome
Peptidoglycan (murein, mucopeptide)
Gram stain
Gram negative
Gram positive
Cell envelope
Cell membrane
Cell wall
Outer membrane
Periplasmic space
Oxidative phosphorylation
Spheroplast/protoplast
Flagella
Chemotaxis
Axial filament
Periplasmic binding protein
Permeases
Storage Granules
Pili (fimbriae)
Capsule (slime layer, glycocalyx)
Endospore
 Figure 1a
The domains of life
Figure 1a
The domains of life
Prokaryotes and eukaryotes
"True" bacteria (which include all bacteria that infect man) are members of one kingdom (the eubacteria, bacteria). In addition, a group of organisms often found in extreme environments form a second kingdom (archaebacteria, Archaea). Morphologically, the two kingdoms of organisms appear similar, especially in the absence of a nucleus, and thus are classified together as prokaryotes. However, they have major biochemical differences. Most archaea live in environments such as hot sulfur springs where they experience temperatures as high as 80 degrees C and a pH of 2. These are called thermoacidophiles. Others live in methane-containing (methanogens) or high salt (extreme halophiles) environments.
Archaea
Based on DNA sequence similarities, it appears that the archaea and eukaryotes diverged from the eubacteria before they diverged from each other (figure 1a) and in some ways, archaea are biochemically more like eukaryotes than they are the eubacteria. For example, the RNA polymerase of archaea is as complex, in terms of number of subunits, as the eukaryote nuclear polymerases and there is considerable amino acid homology with some of the eukaryotic subunits. Gene promoter structure in archaea is also more similar to that of eukaryotes than eubacteria, although, like the eubacteria, archaea have operons and transcribe these to polycistronic mRNA. Similarity also exists between the protein synthesis factors of archaea and eukaryotes suggesting that the overall protein synthesis mechanisms of eukaryotes and archaea may be similar. The 16S rRNAs of the eubacteria and the archaea are quite distinct in sequence.
Eubacteria (with the exception of the genera Mycoplasma and Chlamydia) possess peptidoglycan (synonyms: murein, mucopeptide, cell wall skeleton). Peptidoglycan, contains a unique sugar, muramic acid, not found elsewhere in nature. Archaebacteria contain a pseudomurein that is different in structure from eubacterial murein.
In view of the increasing number of similarities between the archaea and the eukaryotes, the term archaebacteria is no longer used. All other cellular forms of life (including plants, animals, and fungi) are referred to as eukaryotes.
Members of the Archaea are not human pathogens and will not be discussed further.
|
Similarities between Archaea and Eukaryotes |
|||
| Eubacteria | Archaea | Eukaryotes | |
| Nucleus | No | No | Yes: membrane-bound |
| Nucleosomes/histones | No | Yes | Yes |
| Operons/polycistronic mRNAs | Yes | Yes | No |
| Introns | No | No | Yes |
| TATA Box binding protein | No | Yes | Yes |
| Organelles | No | No | Yes: mitochondria, lysosomes, endoplasmic reticulum etc. |
| Chromosomes | One Circular | One Circular | More than one |
| RNA polymerase | One (simple) | More than one (complex) | More than one (complex) |
| Protein initiator amino acid | N-formyl methionine | Methionine | Methionine |
| Protein synthesis sensitivity to diphtheria toxin | Insensitive | Sensitive | Sensitive |
| Peptidoglycan | Yes | No | No |
| Protein synthesis |
of Archaea are more similar to those of eukaryotes than eubacteria |
||
Differences between prokaryotes/eukaryotes
The prokaryotic cell, in contrast to the eukaryotic cell, is not compartmentalized. Nuclear membranes, mitochondria, endoplasmic reticulum, Golgi body, phagosomes and lysosomes are not present (Figures 1b, 2 and 3). Prokaryotes generally possess only a single circular chromosome. Since there is no nuclear membrane, the chromosome is bound to a specific site on the cell membrane - the mesosome. Prokaryotic ribosomes are 70S (S stands for Svedberg unit, a measure of size), whereas eukaryotic ribosomes are larger (80S). Prokaryotic ribosomal subunits are 30S and 50S (eukaryotic are larger). The 30S ribosome has 16S RNA, whilst the 50S ribosome contains 23S and 5S RNA. Ribosomal RNA is larger in eukaryotes (e.g. 18S versus 16S rRNA). Bacterial membranes generally do not contain sterols (e.g. cholesterol).

Figure 1b. Comparison of eukaryotes and "eubacterial" prokaryotes
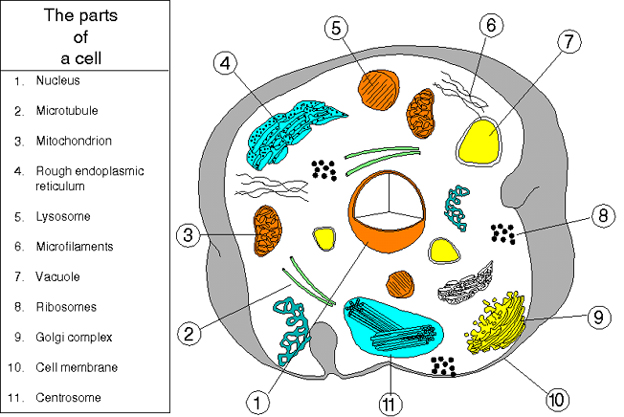
Figure 2. An animal cell
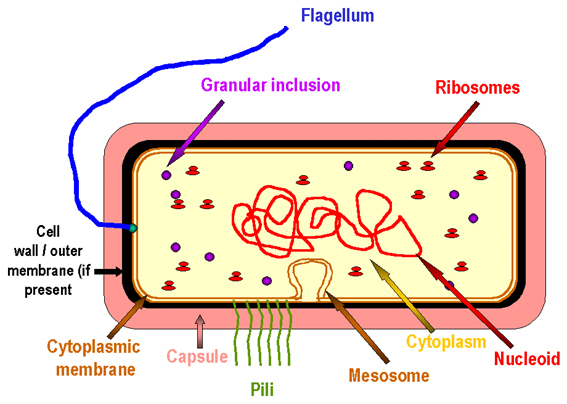
Figure 3. The prototype bacterial cell
BACTERIAL STRUCTURES
Despite their lack of complexity compared to eukaryotes, a number of eubacterial structures may be defined. Not all bacteria possess all of these components.
Plasmids
These are extra-chromosomal DNA, usually present in multiple copies, that often code for pathogenesis factors and antibiotic resistance factors. Some forms are also involved in bacterial replication.The cell envelope
Bacteria can be divided into two groups on the basis of staining with the Gram stain; Gram positive bacteria remain stained by crystal violet on washing, Gram negative do not. All bacteria have a cell membrane where oxidative phosphorylation occurs (since there are no mitochondria). Outside the cell membrane is the cell wall which is rigid and protects the cell from osmotic lysis. In Gram positive bacteria, the cell wall peptidoglycan layer is a much thicker layer than in Gram negative bacteria. Gram negative bacteria have an additional outer membrane. The outer membrane is the major permeability barrier in Gram negative bacteria. The space between the inner and outer membranes is known as the periplasmic space. Gram negative bacteria store degradative enzymes in the periplasmic space. Gram positive bacteria lack a periplasmic space; instead they secrete exoenzymes and perform extracellular digestion. Digestion is needed since large molecules can not readily pass across the outer membrane (if present) or cell membrane.
Wall-less forms of Bacteria
When bacteria are treated with 1) enzymes that are lytic for the cell wall e.g. lysozyme or 2) antibiotics that interfere with biosynthesis of peptidoglycan, wall-less bacteria are often produced. Usually these treatments generate non-viable organisms. Wall-less bacteria that can not replicate are referred to as spheroplasts (when an outer membrane is present) or protoplasts (if an outer membrane is not present). Occasionally wall-less bacteria that can replicate are generated by these treatments (L forms).
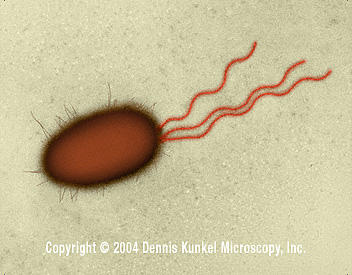
Figure 4. E. coli - rod prokaryote with multiple flagella (SEM x26,730) ©
Dennis Kunkel Microscopy, Inc.
Used with permission
Flagella
Some bacterial species are mobile and possess locomotory organelles - flagella (Figure 4). Those that do are able to taste their environment and respond to specific chemical foodstuffs or toxic materials and move towards or away from them (chemotaxis). Flagella are embedded in the cell membrane, extend through the cell envelope and project as a long strand. Flagella consist of a number of proteins including flagellin. They move the cell by rotating with a propeller like action. Axial filaments in spirochetes have a similar function to flagella. Binding proteins in the periplasmic space or cell membrane bind food sources (such as sugars and amino acids) causing methylation of other cell membrane proteins which in turn affect the movement of the cell by flagella. Permeases are proteins that then transport these foodstuffs through the cell membrane. Energy and carbon sources can then be stored when necessary in cytoplasmic "storage granules" which consist of glycogen, polyhydroxybutyrate or polyphosphate.
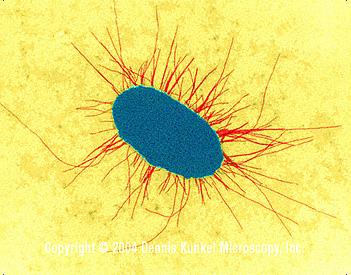
Figure 5. E. coli with fimbriae (TEM x17,250) © Dennis Kunkel Microscopy, Inc. Used with permission
Pili (synonym: fimbriae)
The types of pili (or whether they are produced at all) varies both among and between species. Pili are hair-like projections of the cell (Figure 5). Some are involved in sexual conjugation and others allow adhesion to host epithelial surfaces in infection.
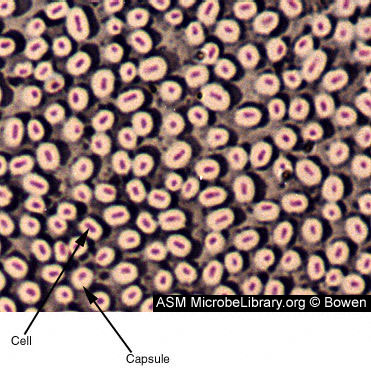 Figure 6
Figure 6Capsule-producing bacillus-shaped bacteria. The capsule is composed of polysaccharides and polyproteins. Capsules have a role in adherence, virulence, protection, securing nutrients, and cell-to-cell recognition. Capsules vary in thickness and can easily be 2 times the volume of the organism. In a capsule stain, the background is stained grayish blue and the cells are stained red. The capsule is unstained and appears as a halo around the cell. © Judy Bowen
Buckman Laboratories International, Inc.
Memphis, Tennessee, USA and The MicrobeLibrary
Capsules and slime layers (Figure 6)
These are structures surrounding the outside of the cell envelope. When more defined, they are referred to as a capsule when less defined as a slime layer or glycocalyx. They usually consist of polysaccharide; however, in certain bacilli they are composed of a polypeptide (polyglutamic acid). They are not essential to cell viability and some strains within a species will produce a capsule, whilst others do not. Capsules of pathogenic bacteria inhibit ingestion and killing by phagocytes. Capsules are often lost during in vitro culture.Endospores (spores)
These are a dormant form of a bacterial cell produced by certain bacteria when starved (figure 7); the actively growing form of the cell is referred to as vegetative. The spore is resistant to adverse conditions (including high temperatures and organic solvents). The spore cytoplasm is dehydrated and contains calcium dipicolinate (dipicolinic acid - figure 8) which is involved in the heat resistance of the spore. Spores are commonly found in the genera Bacillus and Clostridium.
 Figure 7 A
Figure 7 ABacillus cereus spores (green) and cells not forming spores (pink) © Ralph Van Dyke Jr. The Des Moines University Osteopathic Medical Center, Des Moines, Iowa USA and The MicrobeLibrary
 B
B
P. larvae sporulation in culture appears similar to that of other
endospore formers. The rod-shaped vegetative cell has a thick peptidoglycan
layer.
© Donald Stahly, University of Iowa, Iowa City, Iowa
and The MicrobeLibrary
An immature spore is shown surrounded by the mother cell (sporangium). A copy of the bacterial DNA is encased within the developing spore. The outer spore coat appears thinner and less electron dense than in the mature spores. © Donald Stahly, University of Iowa,
Iowa City, Iowa and The MicrobeLibrary
 D
D
The thick spore coat indicates that endospore differentiation is
complete, but the endospore remains within the sporangium. Finally, the
endospore is released from the sporangium. The inner spore coat consists of a
maximum of seven distinct layers referred to as lamellae.
© Donald Stahly, University of Iowa, Iowa City, Iowa
and
The MicrobeLibrary
 Figure 8. Dipicolinic acid
Figure 8. Dipicolinic acid
![]() Return to the Bacteriology Section
of Microbiology and Immunology On-line
Return to the Bacteriology Section
of Microbiology and Immunology On-line
This page last changed on
Friday, February 26, 2016
Page maintained by
Richard Hunt