|

Figura
2
ARNi mund të përdoret direkt nga ARN virusët me varg pozitiv

Figura
3
ARN virusët me varg negativ duhet të krijojnë një varg pozitiv para se
të krijojnë proteinë dhe kështu duhet të paketojnë proteinën polimerazë

Figura
4
Retrovirusët duhet të paketojnë polimerazën e tyre në virion

Figura
5
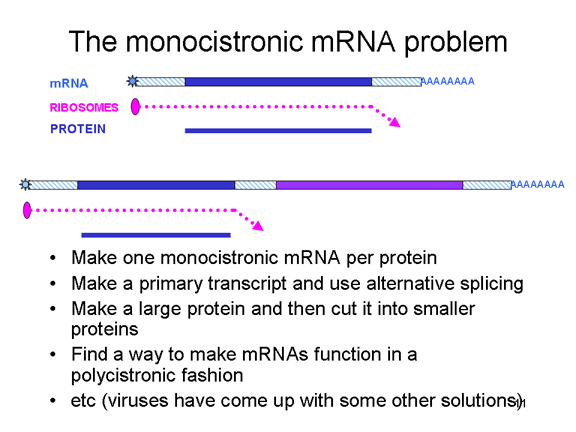
Problemi i ARN monocistronike: zgjidhjet

Figura
6
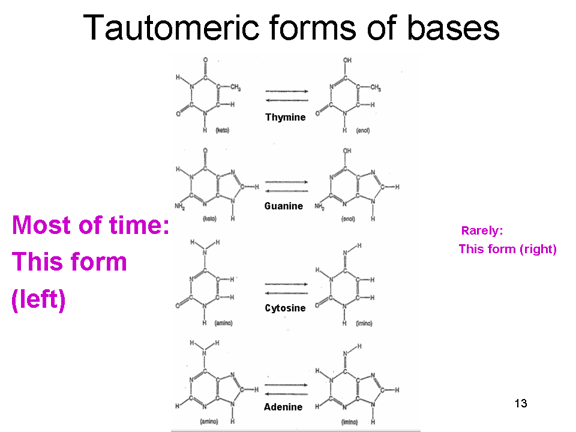
Format tautomerike të bazave të ADN-së

Figura
7
Viruset adaptohen në fitnes në një mjedis të caktuar (virusi i
gjelbërt). Mund të ketë trajta tjera të virusit ( e kuqe) që mund të
jenë po aq të suksesshëm, por për të mbërritur në atë stad , virusi
duhet të kalojë nëpër një formë që nuk mund të garojë në këtë mjedis dhe
si rrjedhojë do të zbutet në popullacion

Figura
8
Pasojat e madhësisë së gjenomit

Figura 9
Poliovirusi
Copyright J.Y. Sgro (përdorur me leje)


Figura
10
Kapja për sipërfaqen e qelizës dhe hyrja e ARN në qelizë
 A
A
 B B
 C
C
 D
D
 E
E
 F
F
|
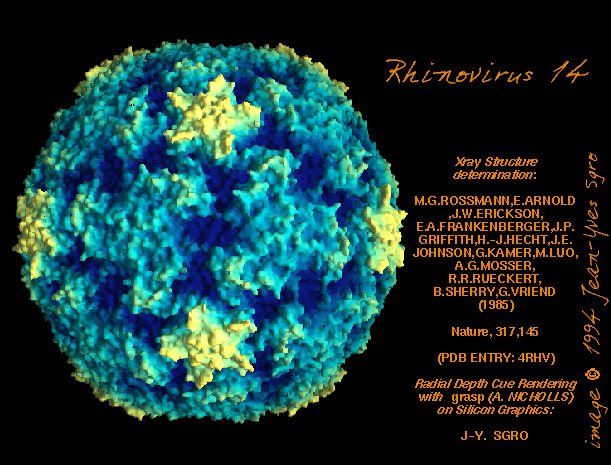
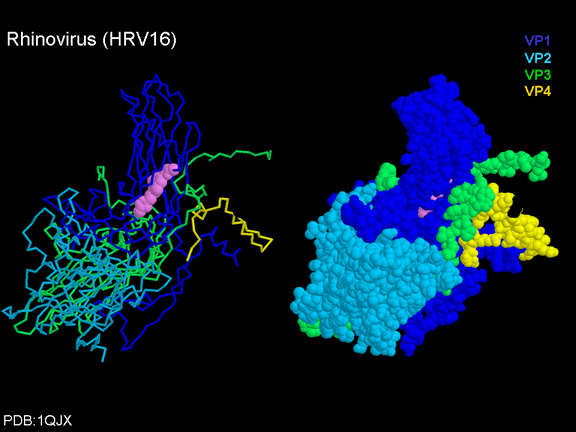
Figura 11 (A)
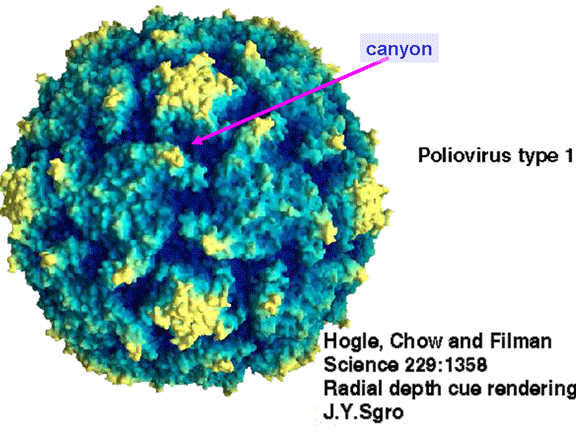
Sipërfaqja e poliovirusit duke e treguar kanionin
Figura
11 (B)
Rinovirusi human 14
Figura
11 (C)
Rinovirusi 14
në
kompleks me receptorët
ICAM-1, e zgjidhur
përmes
rikonstruimit të
pamjes
së krio-elektron
mikroskopisë
© Dr J-Y Sgro,
Universiteti
i Wisconsin.
Përdorur me
autorizim
Figura
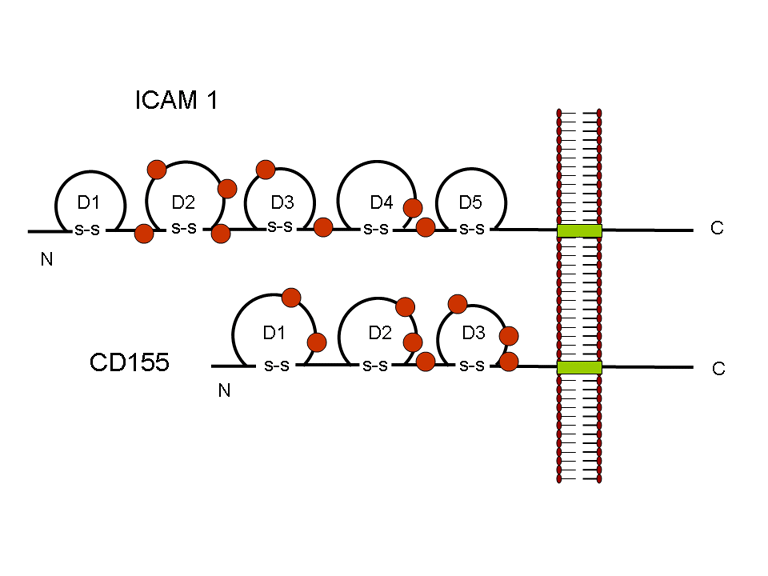
11D
Struktura e regjionit të ICAM1 dhe CD155. ICAM1 ka pesë
regjione ekstracelulare përderisa CD155 ka tre
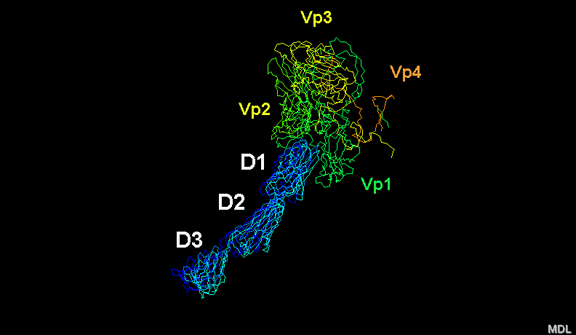
Figura 11E
Receptori i polio virusit (CD155) duke ndërvepruar me
xhepin në virus. Proteinat virale janë VP1, VP2, VP3
dhe VP4
Figura 11F
Rhinovirusi 14 i lidhur për receptorin e tij
© Dr J-Y Sgro, Universiteti i Wisconsin.
Përdorur
me autorizimn |
|
ARN
INFORMATIV I VIRUSIT
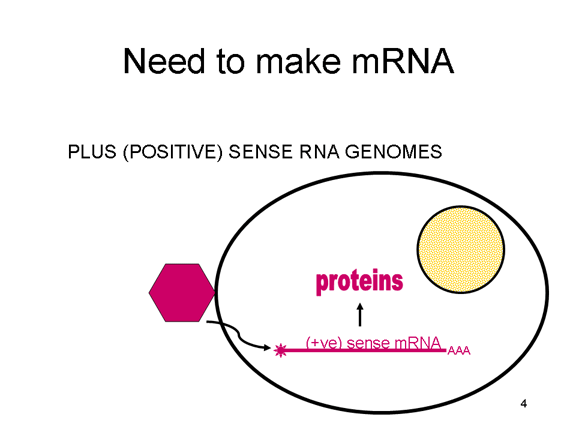
ARN virusi duhet të krijojw një ARN, i cili mund të shërbejw si ARN
informativ për sintezën e proteinave në qelizën e strehuesit. Në rastin
e ARN virusëve me varg pozitiv (gjenomi i të cilëve, për nga definicioni,
ka ngarkesë të njëjtë me ARNi), ARN e gjenomit mund të shërbej si
informatë (figura 2). Kapsida e virusit shërben si mjet dërgues për në
citoplazmë. Pasi që gjenomi është ARN dhe kopjohet nga ARN polimeraza,
nuk ka nevojë për promotor tipik (TATA box, CAT box etj.) që përshkon
për së larti gjenet që kodojnë proteinat (siç ndodh tek ADN virusët ose
retrovirusët); madje, virusët me varg pozitiv nuk kanë nevojë për të
krijuar proteina të reja para se të krijojnë ARNi, pasi që ARN e tyre
mund të shërbej drejtpërdrejt si informatë.
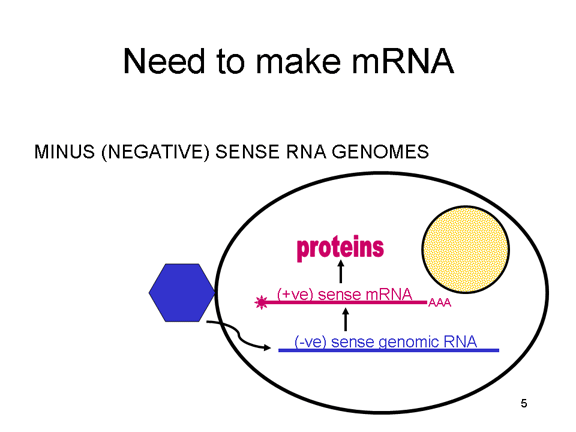
Për dallim, qelizat e infektuara nga virusët me varg negativ, duhet të
përmbajnë proteinën (polimerazën) para se të mund të krijohet ARNi me
varg pozitiv dhe kështu virusi duhet të bart polimerazën brenda kapisdës
dhe ta dërgojë këtë proteinë në qelizën e infektuar (figura 3). Përpos
kësaj, ARNi e kopjuar nga ARN gjenomike negative duhet të ketë
modifikimet normale të ARNi të qelizës (kapela metilike, bishti poli A)
në mënyrë që të përkthehet nga qeliza strehuese. Kjo d.m.th. se enzimat
modifikuese (metilaza, GMP transferaza, Poli A polimeraza) duhet të jenë
të përfshira në virus. Ndonëse qeliza strehuese mund të sigurojë këto
enzime, ato ndodhen në bërthamë (aty ku krijohet ARNi) dhe, siç e kemi
parë më sipër, shumica e ARN viruseve janë citoplazmatik gjatë ciklit të
tyre jetësor.
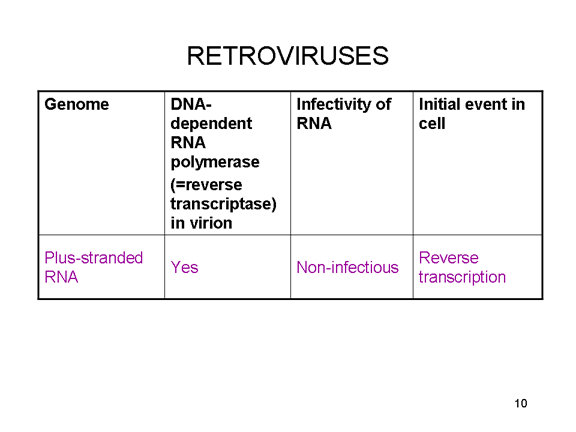
Në rastin e retroviruseve, kopjimi i ARN në AND është i domosdoshëm para
se të krijohet proteina dhe kështu polimeraza (transkriptaza reverse)
duhet gjithashtu të përfshihet në grimcën e virusit (figura 4). Këta ARN
virusë, megjithatë, e replikojnë gjenomin e tyre në bërthamën e qelizës
strehuese dhe kështu mund të përdorin enzimat modifikuese të ARN-së të
qelizave strehuese.
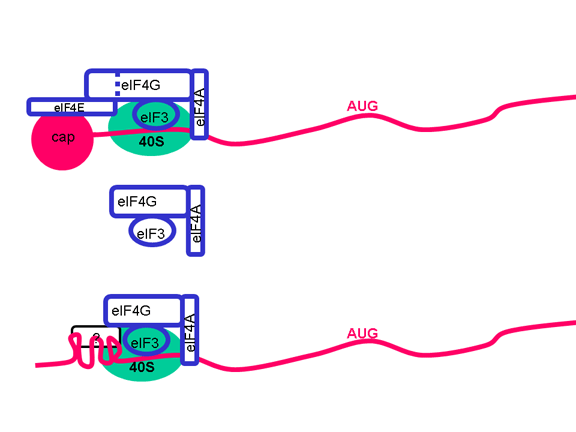
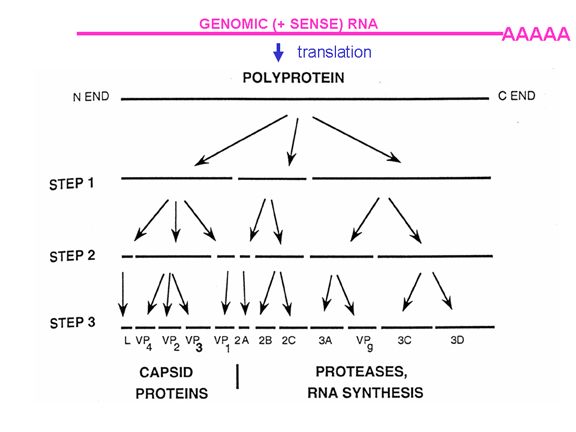
PROBLEMI I ARN
MONOCISTRONIKE
Qelizat eukariotike kanë ARNi monocistronike, për dallim nga bakteret të
cilat mund të kenë ARNi policistronike. ARNi monocistronike e kodon një
proteinë, pasi ribozomi e gjen vendi i inicimit të përkthimit afër
skajit 5’ të ARNi dhe e përkthen proteinën deri sa të arrijnë tek stop
kodoni. Inicimi i brendshëm i sintezës së proteinave brenda ARNi nuk
ndodh (përpos në gjendje të ralla). Një ARN e vetme mund të kodojë më
shumë se një proteinë tek eukariotet por kjo ndodh me anë të ngjitjes së
pjesëve të transkriptimit origjinal për të krijuar një ARNi tjetër, e
cila shërben si informatë monocistronike. Enzimet ngjitëse që e kryejn
këtë proces janë gjetur në bërthamë (pasi këtu krijohet ARNi). Pasi ARN
virusët zakonisht janë citoplazmatik, ato nuk mund të shfrytëzojnë
dobinë e enzimave lidhëse. Prandaj, ARN virusët të cilët kanë vetëm një
ARNi janë të detyruar që të krijojnë vetëm një proteinë të madhe – por
ato kanë zhvilluar një numër të truqeve për tejkaluar këtë (figura 5)
dhe me të vërtetë krijojnë më shumë se një proteinë. Disa mund të
shfrytëzojnë mundësinë e enzimave lidhëse alternative të qelizave
strehuese (prandaj duhet të kenë fazën e bërthamës). Të tjerat e
krijojnë vetëm një proteinë të gjatë të vetme, e cila ka aktivitetin e
proteazës; kjo e pret paraardhësin e gjatë në një seri të proteinave më
të vogla. Të tjerat, siç janë pikornaviruset, kanë gjetur mënyrat për të
krijuar një funksion të vetëm ARNi sipas mënyrës policistronike,
megjithëse janë në qelizë eukariotike Si e bëjnë këtë do të adresohet më
vonë.
PROBLEMI I LEXIMIT TË
DËSHMISË (PROVËS)
Kur ne e kopjojmë ADN gjenomin tonë, përdorim ADN polimerazën e leximit
të dëshmisë, e cila i korrigjon gabimet në kopjen e re të ADN-së.
Shumica e gabimeve ndodhin pasi format normale të bazave të cilat e
përbëjnë kodin në ADN mund të ekzistojnë në forma te rralla tautomerike
(figura 6). Këto forma të rralla ekzistojnë për një kohë shumë të
shkurtë, por nëse baza është në formën e saj tautomerike në kohën kur
kopjohet, do të lexohet si bazë e gabuar gjatë sintezës së ADN-së.
Kështu që, zakonisht A lidhet T por tautomeri i rrallë i T do të
bashkohet në bazë me G dhe do të kopjohet asisoj. ARN polimerazat nuk
kanë zakonisht lexim të dëshmisë, prandaj këto mutacione nuk do të
korrigjohen dhe do të përjetësohen në gjeneratat e ardhshme. Shkalla e
mutacioneve të ARN polimerazës II është afërsisht 1 në 10,000, kështu që,
për të mbajtur nivelin e mutacionit të ARN viruseve të ulët, ato duhet
të kenë gjenome të vogla; zakonisht, ARN virusët kanë gjenome me 10,000
ose më pak nukleotide (në krahasim, p.sh. me me herpes-virusët, të cilët
kanë gjenome prej rreth 150,000 nukleotideve).
PSE TËRË KËTO
MUTACIONE NUK I BËJNE JOSTABIL arN VIRUSËT?
Përgjigja është se i bëjnë jostabil në shumicën e rasteve. Virusi HIV (i
cili ka gjenomin e madhësisë më pak se 10,000 nukleotide) akumulon shumë
mutacione, të cilat rezultojnë me polimorfizëm të kopulacionit dhe i
mundësojnë virusit që të jetë rezistent ndaj barnave antivirale.
Megjithatë, shumë ARN virus (p.sh. poliovirusi) janë shumë stabil
përgjatë mija viteve, pavarësisht nga shkalla e tyre e lartë e
mutacioneve. Kjo mund të shpjegohet më së miri me konceptin e fitnesit
në peisazhin Mullerian. Sipas këtij koncepti, një virus i caktuar mund
të adaptohet lehtë për t’u replikuar në strehuesin e caktuar (p.sh.
polio tek njerëzit). Mund të ekzistojnë forma të tjera të virusit, të
cilat mund të jenë njëlloj ose më të suksesshme, por për të pësuar
mutacion në një formë tjetër virusit do t’i duhej të kalojë nëpër forma
të cilat janë më pak të përshtatura. Kështu që, kur virusi, i cili
adaptohet mirë pëson mutacion, virusët mutantë nuk mund të konkurrojnë
dhe dobësohen. Me fjalë të tjera, virusët mirë të përshtatur kanë një
kulminacion të fitnesit në ambientin e tyre dhe nuk janë në gjendje që
të kalojnë luginën e të përshtaturit të dobët (figura 7). Kjo është
sikur ketri në skajet veriore dhe jugore të Grand Kanjonit, të cilët
kanë divergjuar sepse nuk mund të përshtaten në kushtet e ashpra në
dyshemenë e kanjonit, prandaj edhe nuk mund të shumëzohen.
PASOJAT E MADHËSISË
SË VOGËL TË GJENOMIT
Gjenomi i vogël e kufizon numrin e proteinave të cilat mund të kodohen (figura
8). Është e nevojshme proteina e kapsidës për të mbrojtur ARN-në dhe
polimerazën, e cila duhet të krijohet nga virusi. Gjithashtu, është e
nevojshme të jetë edhe një proteinë për kapje në mënyrë që virusi të
mund të infektojë qelizën e re. Në këtë mënyrë, do të ketë një numër të
kufizuar të proteinave individuale megjithëse ARN virusët mund të jenë
mjaft të mëdhenj duke shfrytëzuar kopje të shumëfishta të proteinave të
ndryshme.
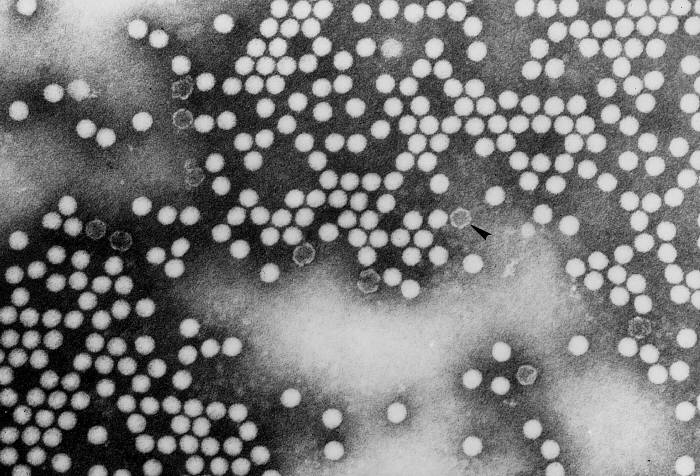
CIKLI JETËSOR
Polioviruset dhe rinovirusët bëjnë pjesë në grupin e pikornavirusëve (figura
9). Ato janë ARN virus të vegjël, ikozaedral, me ngarkesë pozitive të
ARN-së, të cilët nuk kanë mbështjellës (dy shtresa lipidike). Në
pikornaviruse, përfshihen virusi i hepatitit A dhe enterovirusët. Tërë
cikli jetësor i virusit ndodh në citoplazmën e qelizës strehuese.
Meqenëse ARN është i të njëjtës ngarkesë sikur ARNi, ARN-ja e vetme
është infektive, ndonëse e një niveli shumë më të vogël sesa grimca e
plotë e virusit. Kjo ndodh sepse virusi ka proteina, të cilat ngjiten
për sipërfaqe të qelizës.
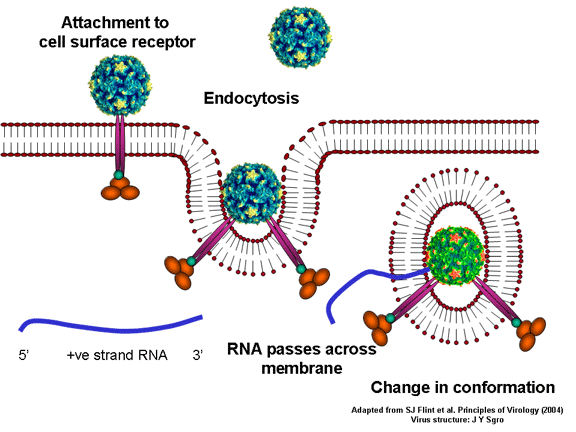
Picornavirusët ngjiten me qelizën strehuese përmes receptorit specifik,
siç bëjnë pothuajse të gjithë virusët dhe internalizohen nga qeliza (figura
10). Në rastin e polio dhe rinoviruseve, kjo është proteinë e një
funksioni të panjohur (në qelizat normale) e njohur si CD155, receptor i
polio virusit RPV. Ekzistojnë disa ide lidhur me funksionin normal të
CD155, në atë që lidhet për proteinën e quajtur vitronektinë, kështu që
mund të jetë një proteinë adezive e përfshirë në lidhjen e qelizës më
matriksin e saj ekstracelular. Shumica e rinovirusëve mund të lidhen
edhe me një receptor tjetër, i cili është një proteinë adezive e quajtur
CD54 ose ICAM-1 (molekula adezive intracelulare-1 – MAIC-1); përdorimi i
këtyre receptorëve e kufizon qelizën të cilën virusi mund ta infektojë,
duke çuar në tropizmin e tyre karakteristik; për shembull, CD155 është i
shprehur në disa lloje të qelizave, duke përfshirë zorrët dhe gjithashtu
edhe në pjesën terminale të aksonit në vendbashkimin neuromuskular, pra
në membranën presinaptike. CD155 dhe ICAM-1 janë glikoproteina, të cilat
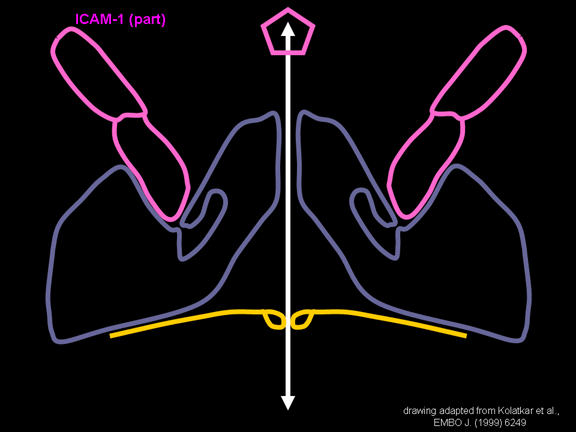
janë të lidhura në membranë dhe të cilën e rrethojnë (figura 11D).
Regjionet e tyre ekstracelulare, për të cilat lidhen pikornaviruset ,
kanë domenet tre dhe katër të ngashme me imunoglobulinat, respektivisht,
me mbështjellës të ngjashëm me imunoglobulinat. Në cilindo rast, virusi
lidhet me hapësirën N-terminale. ICAM-1 është më i gjatë dhe depërton
thellë brenda në kanjon (figura 11 C, D) përderisa CD155 është më i
shkurtër dhe shtrihet përgjatë kanjonit (figura 11E).
Nëse shikoni strukturën e nukleokapsidës do të shihni dymbëdhjetë
pentone një në secilën majë të virusit. Secili penton është në kontakt
me pesë proteina të tjera të cilat e rrethojnë atë. Secila faqe e
virusit ka një kompleks të proteinave, të cilat duken sikur trekëmbësh (strukturë
me tre këmbë). Kjo është në kontakt me gjashtë proteina të tjera (tre
pentone dhe tre trekëmbësha) dhe nganjëherë quhet hekson, ndonëse nuk
duket sikur hekson. Pasi ikozaedri ka njëzet faqe, kemi 20 trekëmbësha.
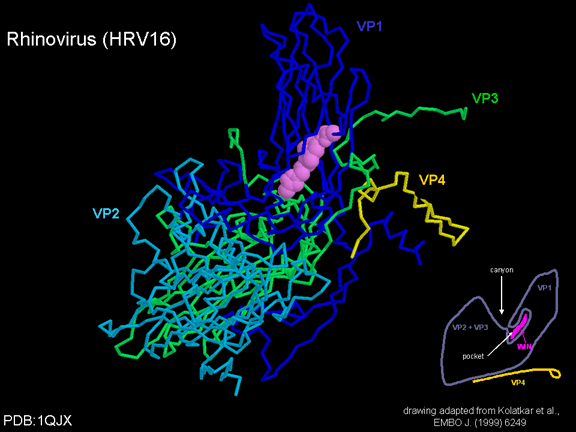
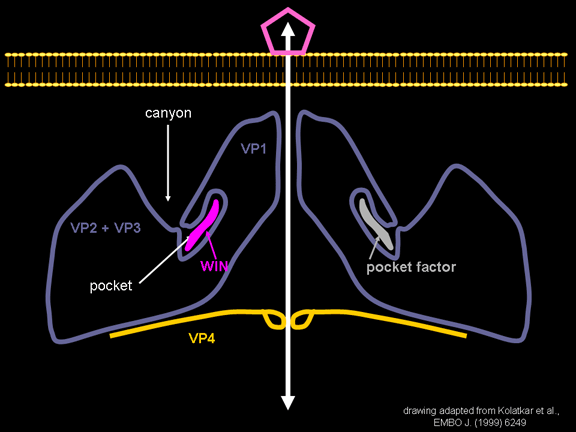
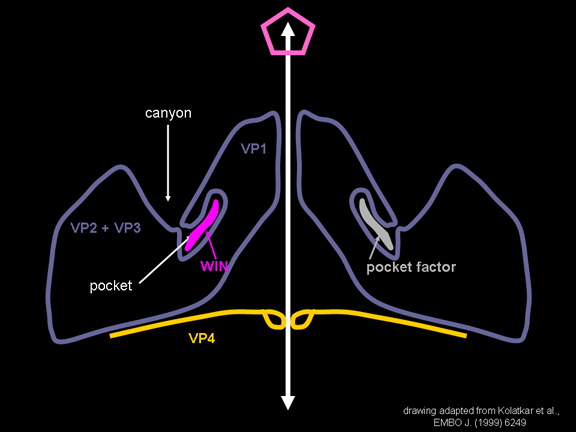
Në mes të pentoneve dhe trekëmbëshave kemi një kanion (figura 11 A, B);
receptori në sipërfaqen e qelizës (ICAM-1 dhe CD155) lidhet në kanion (figura
11E) dhe përshpejton ndryshimin strukturor tek disa prej proteinave
virale. Ndërveprimi i qelizës dhe proteinës virale nuk është aq stabil.
Në rastin e rinovirusëve, hyrja në citoplazmë duket se e përcjell
endocitozën, e cila mund të sugjerojë se është e nevojshme një pH
acidike për ndryshimet strukturore.
Le të shohim më në detaje se çka ndodh kur një rinovirus lidhet për
qelizën përmes ICAM-1. ARN do të hyjë në qelizë nëpër membranë përmes
qendrës së pentonit, pasi që njëra nga proteinat virale ngulitet në
membranën qelizore. Nëse marrim një prerje tërthore nëpër penton (figura
12) (e cila ka simetri pesë shtresore kur shikojmë poshtë në te), mund
të shohim se molekulat VP1 formojnë pjesën qendrore të pentonit dhe e
rrethojnë poren e mbyllur. Kanioni ndodhet aty ku VP1 i përshtatet VP2/3
Por dyshemeja e kanionit nuk është e rrafshët. Ekziston një vrimë e
ngushtë në dysheme, nën të cilën është një hapësirë më e madhe e njohur
si xhep. Normalisht, ky xhep përmban “faktorin e xhepit”. Kjo asnjëherë
nuk është izoluar dhe karakterizuar por duket si një molekulë e gjatë, e
trashë, e ngjashme me acidin në studimet strukturore kristalografike.
Nën proteinat e jashtme, VP1, VP2 dhe VP3 është një proteinë tjetër e
virusit, e cila nuk mund të shihet nga jashtë (si në figurën 12) dhe kjo
është VP4. Figura 13 tregon boshtin e proteinave të pentonit me një
strukturë më të përgjithshme në kuadrat. Figura 14 tregon se si ne
mendojmë se molekula ICAM-1 përputhet në kompleksin VP1/2/3. Vëreni se
përputhet në kanion por nuk e depërton xhepin. Ne e shohim këtë përsëri
në figurën 15 me boshtin e proteinave tani të shënuar.
Një lloj i medikamenteve që është përdorur për të trajtuar infeksionet
me pikornaviruse janë medikamentet WIN. Ato i ndalojnë ndryshimet
strukturore, të cilat ndodhin kur virusi dhe ICAM-1 ndërveprojnë (në
figurën 16 mund të shihni se çka ndodh). Medikamenti WIN e zhvendos
“faktorin e xhepit” dhe përputhet duke u ngjeshur brenda xhepit (figura
17) dhe është pothuajse plotësisht e varrosur siç mund të shihet kur ne
e shikojmë pentonin në një pamje me hapësirë-mbushur (figura 18).
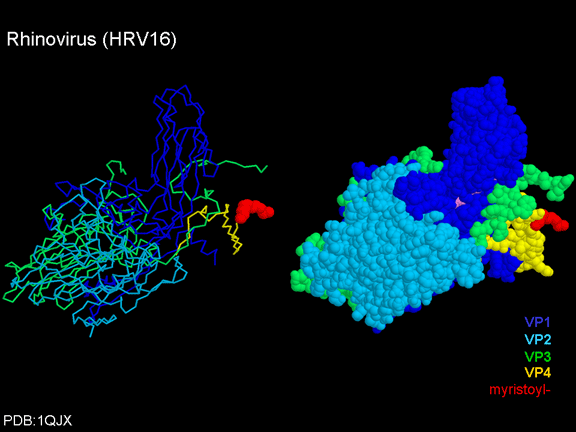
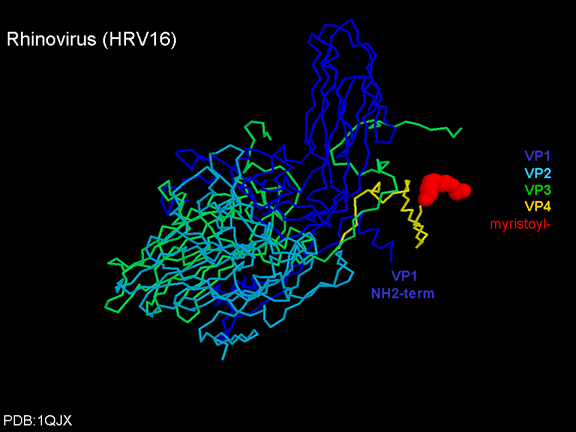
Siç e kemi vërejtur më sipër, një proteinë, VP4, është nën proteinën e
jashtme VP1/2/3. VP4 ka acid yndyror (acidi miristik) të kapur për te (figura
19) e cila e bënë njërin skaj shumë hidrofobik. Në modelin
hapësirë-mbushës (figura 20), ne mund të shohim se acidi yndyror zgjatet
në pore për të formuar një tapë në mesin e pentonit. VP3 gjithashtu e
formon një pjesë të tapës (figura 20).
Kur ICAM-1 lidhet në kanion, kemi një ndryshim strukturor dhe e lëviz
VP1 jashtë qendrës së pentonit (figura 21). Kjo e ngjesh xhepin dhe
është e mundur që lidhja e zhvendos faktorin e xhepit. Tani mund të
shihni pse barërat WIN funksionojnë. Ata lidhen në xhep më fuqishëm sesa
faktori i xhepit dhe nuk zhvendosen me lidhjen e ICAM-1. Kështu që,
përkulja për të hapur poren nuk ndodhë (figura 21b).
Kur VP1 lëvizë mbrapa (figura 21b, c), porja hapet përderisa VP3 ia
lëshon rrugën. Kjo i mundëson VP4 që të lëvizë nëpër pore (kanal). Fundi
aminik i VP4 përmban acid miristik (acid yndyror) dhe kjo në njëfar
mënyre lëvizë nëpër pore. Mbani mend se porja ka simetri pesë shtresore
dhe në të vërtetë janë pesë proteina të tilla VP4 të cilat detyrohen të
kalojnë sipër nëpër kanal. Acidi miristik ngulitet në dyshtresëshin
lipidik të membranës qelizore për të formuar një pore. Formimi i pores
gjithashtu lehtësohet nga fakti se skaji aminik i VP1 është një alfa
heliks amfipatike, e cila rrotullohet jashtë me lidhjen e ICAM-1.
Siç mund të pritej nga struktura e tetramerit VP1-4, VP4 nuk do të duhej
të lidhej nga anti-VP4 antitrupat nga pjesa e jashtme e virusit. Nëse ne
e lëmë virusin për një kohë të gjatë në tretje, VP4 shenjohet sikurse
virusi “frymon”. Kur WIN lidhet në xhep, frymimi i tillë ndalet duke
treguar se lidhja më e tendosur e barit në xhep e stabilizon strukturën
e virusit. Siç mund të pritej, nëse e shtojmë një pjesë të molekulës
ICAM-1, e cila lidhet për kanion, kemi një ndryshim të formës dhe ARN
del jashtë pores dhe virusi nuk është më infektiv. Pentoni nuk është
vetëm vendi i daljes së ARN në qelizën e cila po infektohet, por është
gjithashtu edhe vendi i hyrjes së ARN në nukleokapsidë.
|
 A
A
 B
B
|
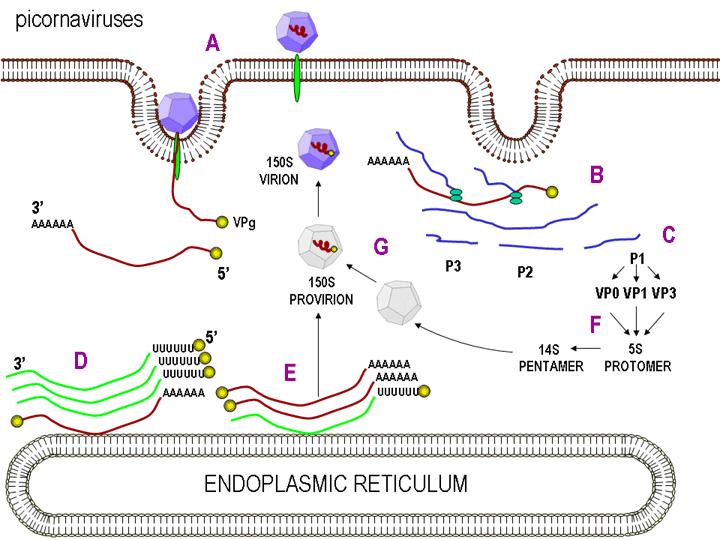
Figura 27
A. Pikornavirusi lidhet për receptorin në sipërfaqen
qelizore (A). ARN me VpG në skajin 5’ përkthehet në një
produkt primar translatues (B) i cili pastaj çahet (C).
ARN gjenomi me varg pozitiv gjithashtu shoqërohet me ARN
polimerazën që është i lidhur për sipërfaqen
citoplazmatike të fshikëzave, me gjasë nga retikulumi
endoplazmatik dhe kopjohet në varg negativ të ARN. VPg
është gjithashtu në skajin 5’ të vargut negativ (skaji
poli U) (D). Vargu negativ kopjohet në ARN gjenomin me
varg pozitiv (E) i cili shoqërohet me prokapsiden për të
formuar 150 S virusin (G), i cili lirohet pas lizës së
qelizës
B. Replikimi i ARN virusit me varg pozitiv |
|
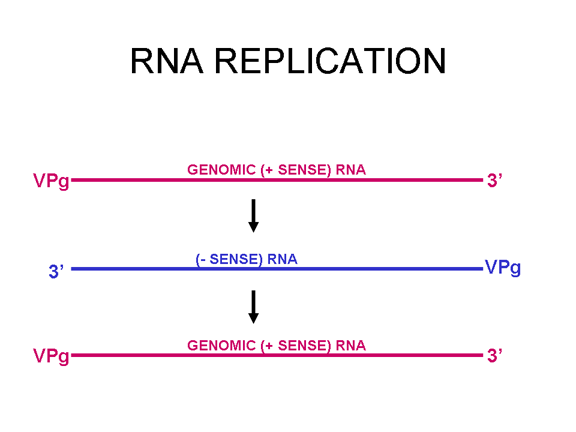
REPLIKIMI I
ARN
Kjo është mjaft e thjeshtë krahasuar me disa ARN virusë të tjerë. Pasi
që pikornavirusët e kalojnë tërë kohën në citoplazmë, atyre iu duhet të
kodojnë një polimerazë (replikazë), e cila është e ndërtuar nga vargu me
ngarkesë të virusit infektues. Polimeraza e kopjon vargun, i cili pastaj
kopjohet prapë në vargun me ngarkesë, i cili është i paketuar në virus (figura
27). Replikimi i ARN duket se ndodh në sipërfaqen citoplazmatike të
vezikulave membranore, për të cilat lidhet ARN polimeraza. Këto vijnë
nga retikulumi endoplazmik, ashtu sikurse vezikulat në qelizën e
painfektuar, të cilat transportojnë proteinat e sekretuara dhe
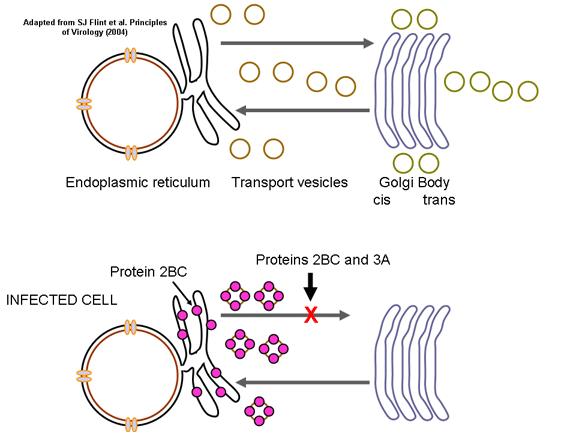
membranore në aparatin e Golgjit. Megjithatë, kur qeliza infektohet nga
pikornavirusi, vezikulat nuk bashkohen me faqen cis të aparatit të
Golgjit, ashtu sikurse bëjnë vezikulat transportuese (figura 28). Këto
vezikula kanë proteina specifike të cakut në sipërfaqet e tyre
citoplazmatike (të quajtura proteinat COP) dhe është e mundur që ato të
jenë të përfshira në përgjigjen ndaj infeksioneve virale. Duhet të
mbahet mend se pikornavirusët nuk kanë mbështjellës lipidik dhe nuk kanë
glikoproteinë sipërfaqësore. Prandaj, prodhimi i virusit nuk inhibohet
me komprometimin e funksionit të aparatit të Golgjit, siç do të ishte
rasti me virusin e mbështjellur. Nuk dihet pse ekziston ky asociacion
membranor i replikimit të ARN-së së pikornavirusit, por mund të
koncentrojë substrate të ndryshme në afërsi të polimerazës (te bakteret,
replikimi i ADN-së është i lidhur me membranën). ARN e pikornavirusëve
është e poliadeniluar në skajin 3’, sikurse janë edhe ARNi qelizore, por
ky poliadenilim ndodh në një mënyrë tjetër. Në sintezën e ARNi në
qelizën strehuese, poli A sekuenca nuk kodohet në kopje të ADN të gjenit
por shtohet nga një enzimë e quajtur poli A polimeraza duke shfrytëzuar
ATP si substrat. Në rastin e ARN-së së pikornavirusit, megjithatë, poli
A sekuenca në vargun me ngarkesë pozitive kopjohet në poli U sekuencë në
skajin 5’ të vargut negativ. Kjo kopjohet përsëri në 3’ poli A përmes
replikazës.
|







 A
A