|
x |
x |
|
 |
 |
|
INFECTIOUS
DISEASE |
BAKTERIOLOGJIA |
IMUNOLOGJIA |
MIKOLOGJIA |
PARAZITOLOGJIA |
VIROLOGJIA |
|
ANGLISHT |
VIROLOGJI –
KAPITULLI XII
NDËRVEPRIMET VIRUS-STREHUES
Dr Gene Mayer
Professor Emeritus
University of South Carolina
Përktheu:
Blerim Berisha
Kryetar i OJQ “Qendra për Avansimin e Studimeve”, Ekspert i
librarisë elektronike biomjekësore në Qendrën e Telemjekësisë së
Kosovës
|
|
En
Español |
|
|
|
|
|
|

 |
|
|
OBJEKTIVAT MËSIMOR
Të përshkruaj mekanizmat e mbrojtjes specifike dhe jospecifike të
strehuesit të përfshira në rezistencë ndaj infeksioneve me virus dhe
shërimin ndaj tyre
Të diskutojë rolin e interferoneve në infeksionet virale
Të rishikojë mekanizmat përmes të cilëve interferoni e shfaq aktivitetin
e tij antiviral
Të diskutojë kontributin relativ të mekanizmave të ndryshëm mbrojtës të
strehuesit në infeksionet virale |
Qëndrueshmëria dhe shërimi nga infeksionet virale varen
nga ndërveprimi që ndodh ndërmjet virusit dhe strehuesit. Mbrojtja, e
cila krijohet
nga strehuesi, mund të veproj drejtpërdrejt në virus ose tërthorazi në
riprodhimin e virusit duke dëmtuar ose mbytur qelizën e infektuar.
Mbrojtja jospecifike e strehuesit funksionon në fazën e kontaktit të
hershëm me virusin, për të parandaluar ose kufizuar infeksionin. Kurse,
mbrojtja specifike vepron pas infeksionit në imunitetin rikuperues ndaj
sfidave pasuese. Mekanizmat mbrojtës të strehuesit, të përfshirë në një
infeksion të caktuar viral do të varen nga virusi, doza dhe porta e
hyrjes së tij. Në vijim janë dhënë disa parime të përgjithshme të
ndërveprimit virus-strehues.
MBROJTJA NDAJ INFEKSIONIT
Mbrojtja e
lindur
Strehuesi i ka disa mekanizma mbrojtës ndaj
infeksionit, të cilët janë të bashkëlindur për organizmin. Këto
paraqesin vijën e parë të mbrojtjes dhe funksionojnë duke e
parandaluar ose kufizuar infeksionin.
Lëkura
Lëkura vepron si barrierë e pamposhtur ndaj shumë
viruseve dhe vetëm pasi kjo barrierë të tejkalohet, virusi do të
mund të infektojë strehuesin.
Mungesa e receptorëve të membranës
Virusët hyjnë në qelizat e strehuesit duke u
lidhur së pari me receptorët specifikë në qeliza (Tabela 1;
adaptuar nga: Roitt, Immunology, 5th ed).
|
|
FJALËT KYÇE
Mbrojtja e lindur
Mbrojtja e nxitur
Interferoni
2'5' Oligo A sintetaza
Protein-kinaza e aktivizuar nga IFN
Aktiviteti i brendshëm antiviral
Aktiviteti i jashtëm antiviral
ADCC
Imunoadherenca
Qelizat VN
LTC-të |
|
|
Tabela 1 |
|
Virusi |
Receptori |
Tipi i qelizës së
infektuar |
|
HIV |
CD4 |
Limfocitet
TH |
|
Virusi Epshtajn-Barr |
CR2 (receptori i komplementit
tip 2) |
Limfocitet B |
|
Virusi i gripit A |
Glycophorin A |
Shumë tipe
qelizore |
|
Rinoviruset |
ICAM-1 |
Shumë tipe
qelizore |
Rangu i strehuesit të virusit do të varet nga
prania e këtyre receptorëve. Pra, nëse strehuesit i mungon
receptori për virusin ose nëse qelizës strehuese i mungojnë disa
komponenta të nevojshme për riprodhimin e virusit, atëherë
strehuesi do të jetë rezistent ndaj atij virusi. P.sh. minjëve
iu mungon receptori për virusin e polios dhe si rrjedhim ata
janë rezistent ndaj poliomielitit. Ngjashëm, njerëzit kanë
rezistencë të lindur ndaj shumë viruseve të bimëve dhe kafshëve.
Mukusi
Mukusi, që e mbulon indin epitelial, vepron si barrierë për të
parandaluar infektimin e qelizës së strehuesit. Në disa raste,
mukusi vepron thjesht si një barrierë, kurse në raste të tjera
mukusi mund të parandalojë infeksionin duke konkurruar me
receptorët e viruseve në qeliza. P.sh. familjet e orthomyxo dhe
paramyxoviruseve infektojnë qelizat e strehuesit duke u lidhur
me receptorët e acidit sialik. Prandaj, glikoproteinat në mukus,
të cilat përmbajnë acidin sialik, mund të konkurrojnë me qelizat
e receptorëve dhe të dobësojnë ose parandalojnë lidhjen e
virusit me qelizën.
Epiteli me cilie
Epiteli ciliar mund t’i ndihmojë strehuesit në dobësimin e
infektivitetit të disa viruseve. Ky sistem është treguar i
rëndësishëm në infeksionet respiratore, meqenëse kur aktiviteti
i këtij sistemi frenohet nga barnat ose infeksionet, kemi një
rritje të shkallës së infektivitetit nga një inokulum i caktuar
i virusit.
pH e ulët
Vlera e ulët e pH në sekrecionet gastrike e inaktivizon shumicën
e viruseve. Megjithatë, enterovirusët janë rezistent ndaj
sekrecioneve gastrike, prandaj mund të mbijetojnë dhe të
shumëzohen në zorrë.
Komponentat humorale dhe qelizore
Shih më poshtë.
|
| |
Barrierat e
nxitura
Ndryshimet të cilat ndodhin tek strehuesi si
përgjigje ndaj infeksioneve, po ashtu, mund të ndihmojnë në
dobësimin e infektivitetit të virusit.
Ethet
Ethet mund të ndihmojnë në frenimin e riprodhimit
të virusit duke fuqizuar mbrojtjet e tjera imunitare dhe duke
zvogëluar riprodhimin e virusit. Riprodhimi i disa viruseve
zvogëlohet në temperaturat mbi 37°C.
pH e ulët
pH e infiltrateve inflamatore është gjithashtu e
ulët dhe mund të ndihmojë në kufizimin e përhapjes së
infeksioneve virale duke i inaktivizuar virusët.
Komponentat humorale dhe qelizore
Shih më poshtë.
KOMPONENTAT
HUMORALE TË PËRFSHIRA NË REZISTENCËN NDAJ INFEKSIONEVE VIRALE
Jospecifike
Në rezistencën ndaj infeksioneve virale funksionon
një numër i komponentave humorale të sistemit imunitar jospecifik.
Disa nga këto komponenta janë të pranishme në mënyrë konstitutive,
përderisa disa të tjera nxiten nga prania e infeksionit.
Interferoni
(IFN)
IFN është zbuluar para më shumë se 40 viteve nga Issacs dhe
Lindemann, të cilët kanë treguar se fraksionet supernatante nga
qelizat e infektuara me virus përmbanin një proteinë, e cila
mund të jep rezistencë ndaj infeksionit për qelizat e tjera. Kjo
substancë nuk vepronte drejtpërdrejt në virus, por vepronte në
qeliza duke i bërë ato rezistente ndaj infeksionit (figura 1).
|
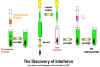 Figura
1 Zbulimi i interferonit
Figura
1 Zbulimi i interferonit
 Figura
2. Përgjigja tipike ndaj infeksionit akut viral
Figura
2. Përgjigja tipike ndaj infeksionit akut viral |
IFN është njëra nga vijat e
para të mbrojtjes kundër viruseve sepse është e nxitur herët
pas infeksionit me virus- para se të paraqiten mekanizmat
tjerë mbrojtës (p.sh. antitrupat, qelizat Tc etj.) (figura
2). Koha, pas së cilës fillon të krijohet IFN, varet nga
doza e virusit.
a) Tipet dhe karakteristikat e interferoneve
Tabela 2; Adaptuar nga: Murray, Medical Microbiology, 5th
ed. tabela 14-3)
|
Tabela 2
Tipet dhe karakteristikat e interferoneve |
|
|
Interferoni |
|
Karakteristika
|
Alfa |
Beta |
Gama |
|
Emërtimi i mëparshëm |
IFN Leukocitar
Tipi I |
IFN Fibrobllastik
Tipi I |
IFN Imunitar
Tipi II |
|
Gjenet
|
>20 |
1 |
1 |
|
Stabiliteti
i pH2
|
Stabil |
Stabil |
Labil |
|
Nxitësit
|
(ARN>ADN)
dvARN Virusët
|
(ARN>ADN)
dvARN Virusët
|
Antigjenet, Mitogjenet |
|
Burimi
kryesor |
Leukocitet, Epiteli |
Fibroblastet |
Limfocitet |
Ekzistojnë tri
tipe të interferonit, IFN-alfa (i njohur edhe si interferoni
leukocitar), IFN-beta (interferoni fibroblastik) dhe
IFN-gama (interferoni imunitar). IFN-alfa dhe IFN-beta quhen
edhe si Tip I, kurse IFN-gama si Tip II. Ekzistojnë rreth 20
nëntipe të IFN-alfa por vetëm një IFN-beta dhe IFN-gama.
Interferonet kanë karakteristika të ndryshme të cilat
shfrytëzohen për t’i dalluar ato (p.sh. stabiliteti dhe
aktiviteti i pH në prani të SDS) por aktualisht
identifikohen duke përdorur antitrupa specifik ndaj
interferoneve.
b) Nxitësit e interferoneve – Qelizat normale nuk përmbajnë
IFN të paraformuar si dhe nuk sekretojnë në mënyrë
konstitutive. Kjo vjen nga fakti se gjenet e interferonit
nuk transkriptohen në qelizat normale. Transkripcioni i
gjeneve të IFN ndodh vetëm pas ekspozimit të qelizave ndaj
nxitësit të duhur. Nxitësit e IFN-alfa dhe IFN-beta
përfshijnë infeksionet me virus, ARN dy vargor (p.sh. poli
inosinik: acidi policitidilik; [poli I:C]), LPS dhe
komponentet nga disa baktere. Nga viruset, ARN virusët janë
nxitësit më të mirë, përderisa ADN virusët janë nxitës të
dobët, me përjashtim të poxvirusëve. Nxitësit e IFN-gama
përfshijnë mitogjenet dhe antigjenet (d.m.th. ato që i
aktivizojnë limfocitet).
|
 Figura 3
Figura 3
Mënyra
e veprimit të interferonit |
c) Ndodhitë qelizore në
nxitjen e interferoneve
IFN gjenet nuk shprehen në qelizat normale sepse qelizat
prodhojnë një proteinë labile represore, e cila lidhet me
regjionin e promotorit mësipër gjenit dhe e inhibon
transkripcionin. Si shtesë, transkripcioni i gjeneve ka
nevojë për proteinën aktivizuese për t’u lidhur me regjionin
e promotorit dhe për të kyçur transkripcionin. Nxitësit e
IFN veprojnë ose duke parandaluar sintezën e proteinës
represore ose duke e rritur nivelin e proteinave aktivizuese
dhe në këtë mënyrë duke i kyçur IFN gjenet. Pasi të largohet
nxitësi, IFN gjeni përsëri çkyçet nga proteina represore dhe/ose
nga mungesa e proteinës aktivizuese. Pasi të aktivizohet
gjeni, ai transkriptohet, ARNi përkthehet dhe proteina
sekretohet nga qeliza. IFN do të lidhet për IFN receptorët
në qelizat fqinje dhe nxisin gjendjen antivirale në qelizën
tjetër (figura 3).
|
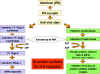 Figura
4
Figura
4
Baza
molekulare e gjendjes antivirale |
d) Ndodhitë qelizore në
veprimin e interferoneve – Lidhja e IFN me receptorin e tij
rezulton me transkripcionin e një grupi të gjeneve, të cilët
kodojnë proteinat antivirale të përfshira në parandalimin e
riprodhimit të viruseve në atë qelizë. Si rrjedhim, qeliza
do të mbrohet nga infeksioni me virus derisa proteinat
antivirale të degradohen (ky proces zgjat disa ditë).
Gjendja antivirale në qelizat e trajtuara me IFN rezulton
nga sinteza e dy enzimeve, të cilat bëjnë inhibimin e
sintezës së proteinave. Njëra proteinë në mënyrë indirekte
ndikon në sintezën e proteinave duke e zbërthyer ARNi viral,
kurse tjetra në mënyrë direkte ndikon në sintezën e
proteinave duke inhibuar zgjatjen (figura 4). Një proteinë e
quajtur 2'5'Oligo A sintetaza, është enzimë e cila e
konverton ATP-në në polimer unik (2'5' Oligo A) që përmban
lidhje 2'- 5'fosfodiesterike. ARN dy vargor është i
nevojshëm për veprimin e këtij enzimi. 2'5'Oligo A pastaj
aktivizon ARNazen L e cila e zbërthen ARNi virale. Proteina
e dytë është proteinë-kinazë, e cila në prani të ARN
dyvargore, autofosforilohet dhe në këtë mënyrë aktivizohet.
Proteinë-kinaza e aktivizuar pastaj fosforilon faktorin
zgjatës eIF-2 dhe e inaktivizon atë. Nga veprimi i këtyre dy
enzimeve të nxitura nga IFN sinteza e proteinave inhibohet.
Megjithëse qeliza e infektuar mund të shkatërrohet si pasojë
e inhibimit të sintezës së proteinave të strehuesit,
progredimi i infeksionit ndalet. Qelizat e painfektuara nuk
vriten nga veprimi i IFN pasi për aktivizimin e dy enzimave
nevojitet ARN dyvargore, e cila nuk prodhohet. Disa viruse
kanë mënyra për të inhibuar efektin antiviral të IFN. P.sh.
adenovirusët prodhojnë ARN, i cili parandalon aktivizimin e
proteinëkinazës nga ARN dyvargore dhe në këtë mënyrë e
zvogëlojnë efektin antiviral të IFN.
|
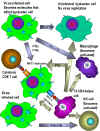 Figura
5
Figura
5
Efektet e interferonit alfa, beta dhe gama |
e) Veprimet tjera biologjike
të interferoneve - IFN jo vetëm që nxit prodhimin e
proteinave antivirale, por gjithashtu ka edhe efekte të
tjera tek qelizat, disa nga të cilat në mënyrë indirekte
kontribuojnë në aftësinë e strehuesit që të rezistojë ose
rikuperohet nga infeksionet virale (figura 5). IFN mund të
ndihmojë modulimin e përgjigjeve imunologjike përmes efektit
të tij në MHC molekulat e klasës I dhe II. IFN-alfa, IFN-beta
dhe IFN-gama e rrisin ekspresionin e molekulave të klasës I
në të gjitha qelizat, duke ndihmuar në njohjen nga
limfocitet Tc, të cilat mund të shkatërrojnë qelizat e
infektuara me viruse. IFN-gama, gjithashtu mund të rrisë
ekspresionin e molekulave MHC të Klasës II, të cilat
përmbajnë antigjenin. Kjo rezulton me prezantimin më të mirë
të antigjeneve virale ndaj limfociteve T ndihmëse CD4+ .
Gjithashtu, IFN-gama mund të aktivizojë qelizat NV, të cilat
mund të vrasin qelizat e infektuara me virus. IFN-të
gjithashtu e aktivizojnë edhe veprimin e brendshëm dhe të
jashtëm antiviral të makrofagjeve për të shkatërruar qelizat
e tjera të infektuara me virus. IFN-të gjithashtu kanë edhe
veprim antiproliferativ, prandaj përdoren në trajtimin e
disa sëmundjeve malinje.
|
| |
f) Përdorimet klinike të
interferoneve – IFN janë përdorur në trajtimin e disa
sëmundjeve virale dhe sëmundjeve të tjera (Tabela 3;
Adaptuar nga: Mims, Medical Microbiology, Fig. 37.5)
|
Tabela 3
Përdorimi klinik i interferoneve |
|
Interferoni |
Përdorimi terapeutik |
|
IFN-alfa, IFN-beta |
Hepatiti
B (kronik)
Hepatiti C
Herpes zosteri
Papilloma virusi
Rinovirusi (vetëm profilaktik)
Lythat |
|
IFN-gama |
Lepra
lepromatoze
Leishmaniaza
Toksoplazmoza
Sëmundjet kronike granulomatoze (SKG) |
|
| |
Përveç kësaj, për shkak të efektit të tyre antiproliferativ,
IFN-të përdoren edhe në trajtimin e shumë sëmundjeve malinje
(Tabela 4; Adaptuar nga: Zinsser, Microbiology, 20th Ed,
Tabela 58.3).
|
Tabela 4
Përdorimi i interferoneve në trajtimin e kancerit |
|
Tumori |
Përqindja e
kompletimit ose remisioni i pjesërishëm |
|
Hairy cell leukemia |
90 |
|
Leukemia
mielocitare kronike |
90 |
|
Limfoma e
limfociteve T |
53 |
|
Sarkoma e
Kaposit |
42 |
|
Neoplazmat
pankreatike endokrine |
30 |
|
Limfomat non-Hoxhkin |
25 - 35 |
|
| |
Megjithatë, efektet anësore të terapisë me IFN e kufizojnë
përdorimin e tyre rutinor në praktikën klinike (Tabela 5;
Adaptuar nga: Mims, Medical Microbiology, Fig. 37.6).
|
Tabela 5
Efektët
më të shpeshta anësore të interferoneve |
|
Interferonet |
Ethet
Këputje
Plogështi
Dhembja e muskujve (mialgjia)
Toksiciteti ndaj:
veshkave
mëlçisë
palcës së eshtrave
zemrës |
|
| |
Komplementi
Shumica e virusëve nuk e fiksojnë komplementin përmes rrugës
alternative. Megjithatë, ndërveprimi i antitrupit
komplement-fiksues me qelizën e infektuar me virus tek viruset
me mbështjellës mund të rezultojë në lizën e qelizës ose të
virusit. Prandaj, në ndërveprim me sistemin imunologik specifik,
komplementi, gjithashtu luan një rol me rëndësi në rezistencën
ndaj infeksioneve virale.
Citokinet
Përveç IFN, edhe citokinet mund të luajn një rol me rëndësi në
rezistencën ndaj infeksioneve virale. Faktori i nekrozës
tumorale alfa (FNT- α), interleukina-1 (IL-1) dhe IL-6 janë
treguar të kenë veprime antivirale in vitro. Këto citokine
prodhohen nga makrofagët e aktivizuar, por kontributi i tyre
ndaj rezistencës in vivo nuk është sqaruar plotësisht.
|
| |
Specifik
Antitrupi i prodhuar nga sistemi imunologjik specifik
është i përfshirë kryesisht në rikuperimin nga infeksionet virale
dhe në rezistencën ndaj sfidave pasuese me virusin. Antitrupat IgG,
IgM dhe IgA mund të luajnë rol në imunitetin ndaj infeksioneve
virale, por kontributi relativ ndaj klasave të ndryshme varet nga
virusi dhe porta e hyrjes. Për shembull, IgA do të jetë më e
rëndësishme tek virusët të cilët infektojnë mukozën; përderisa
antitrupat IgG do të jenë më të rëndësishëm në infeksionet, në të
cilat viremia është veçoria më e theksuar. Antitrupat mund të kenë
efekte edhe të dobishme edhe të dëmshme për strehuesin.
Efektet e
dobishme (Tabela 6: Adaptuar nga: Roitt, Immunology 5th Ed.,
Fig 16.5)
Antitrupi mund të neutralizojë në mënyrë direkte infektivitetin
e virusit duke e penguar lidhjen e virusit me receptorët në
qelizat e strehuesit ose hyrjen e virusit në qelizë. Antitrupat,
gjithashtu mund të pengojnë heqjen e mbështjellësit duke
ndërhyrë me ndërveprimin e proteinave virale të përfshira në
heqjen e mbështjellësit. Antitrupat fiksues të komplementit mund
të asistojnë në lizën e qelizës së infektuar me virus ose të
viruseve me mbështjellës. Antitrupat, gjithashtu mund të
veprojnë edhe si opsonine dhe të fuqizojnë fagocitozën e
viruseve ose duke lehtësuar kapjen e tyre përmes receptorëve Fc
ose C3b ose duke aglutinuar virusët për t’i bërë ato me lehtë të
fagocitueshëm. Qeliza e infektuar me virus, e cila është e
mbështjellur me antitrupa mund të vritet nga qelizat K, me
ç’rast parandalohet përhapja e infeksionit.
|
Tabela 6
Efektet antivirale të antitrupave |
|
Caku |
Agjenti |
Mekanizmi |
|
Virusi i lirë |
Antitrupi i vetëm |
Bllokon lidhjen me qelizë
Bllokon hyrjen në qelizë
Bllokon heqjen e mbështjellësit të virusit |
|
Antitrupi + Komplementi |
Dëmtimi i mbështjellësit
të virusit
Opsonizimi i virusit |
|
Qeliza e infektuar me virus |
Antitrupi + Komplementi |
Liza e qelizës së
infektuar
Opsonizimi i qelizës së infektuar |
|
Antitrupi i lidhur për qelizën e infektuar |
ADCC nga K qelizat, NV
qelizat dhe/ose makrofagët |
Efektet e dëmshme
a) Dëmtimi
imunopatologjik – Fiksimi i komplementit nga imunokomplekset
mund të rezultojë me lirimin e amineve vazoaktive,
rekrutimin e qelizave antiinflamatore dhe dëmtimin pasues të
indit të strehuesit. Disa viruse, siç është virusi i
koriomeningjitit limfocitar prodhojnë sasi të mëdha të
imunokomplekseve në qarkullim, të cilat depozitohen në
shtratin vaskular dhe në veshka, ku e fiksojnë komplementin
dhe rezultojnë me dëmtimin e indit. Shembuj të tjerë të
viruseve të cilët shkaktojnë efekte të tilla janë: morbili,
virusi sincicial respirator, virusi i etheve tropikale dhe
virusi i hepatitit B.
b) Imunoad’herenca – Opsonizimi i viruseve me antitrupin
mund të rrisë kapjen nga qelizat fagocitare. Nëse virusi
është në gjendje të mbijetojë në fagocit, kjo mundëson
përhapjen e infeksionit viral. Ethet tropikale dhe HIV janë
shembuj të viruseve të cilët mund të mbijetojnë në makrofag.
Serologjia
Pasqyra klinike dhe serologjia luajnë një rol të më të madh në
diagnostikimin e sëmundjeve virale, pasi që metodat e tjera (izolimi
dhe identifikimi i viruseve) zakonisht nuk bëhet në mënyrë
rutinore në laboratoret klinike. Tipet kryesore të antitrupave
të cilët kërkohen në diagnostikë janë antitrupat neutralizues,
hemaglutinues, inhibues dhe fiksues të komplementit. Antitrupat
fiksues të komplementit e përcjellin kinetikën e IgM dhe janë më
të dobishëm për të dëshmuar infeksionin e tanishëm ose të
sapokaluar. Për dallim, antitrupat neutralizues dhe
hemaglutinues e përcjellin kinetikën e IgG, perzistojnë për kohë
më të gjatë dhe përdoren për të vlerësuar imunitetin. Krijimi i
antitrupave ndaj komponentave të ndryshme të virusit
shfrytëzohet në etapizimin e sëmundjes. P.sh., kjo përdoret tek
infeksioni me Hepatit B dhe HIV.
|
| |
KOMPONENTAT
QELIZORE
Përveç barrierave dhe komponentave humorale të përfshira
në rezistencë dhe rikuperim nga infeksionet virale, ekzistojnë edhe disa
qeliza të ndryshme të cilat luajnë rol në mbrojtjen tonë kundër viruseve.
Jospecifike
Makrofagët
Makrofagët, përmes mundësisë së ndodhjes në pjesë të ndryshme të
trupit, janë ndër qelizat e para që takohen me viruset. Dëshmitë
eksperimentale sugjerojnë se këto qeliza luajnë një rol të
rëndësishëm në rezistencën ndaj infeksioneve virale. P.sh. miu i
porsalindur është i ndjeshëm ndaj infeksioneve me virusin e
herpesit tip 1, për shkak të defektit në aftësinë e makrofagut
për të penguar riprodhimin e virusit. Mirëpo, makrofagët nga
mijtë e rritur janë në gjendje që të pengojnë riprodhimin e
virusit dhe këta minj janë rezistent ndaj infeksionit me këtë
virus. Gjithashtu, kafshët tek të cilët makrofagët janë në numër
më të vogël janë më të ndjeshëm ndaj infeksionit nga një mori e
viruseve. Makrofagët kontribuojnë në mbrojtjen antivirale në
disa mënyra:
a)
Aktiviteti i brendshëm antiviral – Makrofagët mund të
infektohen me virus por shumë virus nuk janë në gjendje që
të replikohen në makrofag. Makrofagët e aktivizuar (p.sh.
përmes IFN-γ) janë edhe më shumë në gjendje që t’i
rezistojnë replikimin viral. Kështu që, makrofagët ndihmojnë
në kufizimin e infeksionit viral përmes vetisë së tyre të
brendshme që të pengojnë riprodhimin e viruseve. Megjithatë,
disa virus janë në gjendje që të riprodhohen ose së paku që
të mbijetojnë në makrofagë dhe kështu mund të shpërndahen
përmes makrofagut (shih më sipër).
b) Aktiviteti i brendshëm
antiviral – Makrofagët janë gjithashtu në gjendje që të
njohin qelizat e infektuara me virus dhe t’i shkatërrojnë
ato. Makrofagët gjithashtu kontribuojnë në mbrojtjen
antivirale përmes vetisë së aktivitetit citotoksik të tyre.
c) ADCC – Qelizat e infektuara me virus, të cilat janë të
mbështjella me antitrupa IgG mund të shkatërrohen nga
makrofagët përmes ADCC.
d) Prodhimi i IFN – Makrofagët janë burim i IFN.
Qelizat VN
Dëshmitë eksperimentale gjithashtu sugjerojnë se qelizat vrasëse
natyrore(VN) luajnë rol në rezistencën ndaj infeksioneve virale.
Minjët tek të cilët është zvogëluar numri i qelizave VN janë më
të ndjeshëm ndaj infeksioneve me disa virus. Gjithashtu,
pacientët me aktivitet të zvogëluar të qelizave VN janë më të
ndjeshëm ndaj rishfaqjes së infeksionit me virusin Herpes
simpleks tip 1. Qelizat VN veprojnë përmes njohjes dhe vrasjes
së qelizave të infektuara me virus. Njohja e qelizave të
infektuara me virus nuk është e kufizuar nga MHC ose antigjenet
specifike. Prandaj, qelizat VN do të vrasin qelizat e infektuara
nga shumë virus të ndryshëm. Qelizat VN mund të ndërmjetësojnë
ADCC dhe mund të vrasin qelizat e infektuara me virus përmes
këtij mekanizmi. Veprimet e qelizave VN mund të fuqizohen përmes
IFN-γ dhe Il-2 (shih më sipër).
|
| |
Specifike
Limfocitet T – limfocitet T luajnë rol të rëndësishëm në rikuperimin
nga infeksionet virale. Qelizat T citotoksike (LTC-të) të prodhuara
si përgjigje ndaj antigjenit viral në qelizën e infektuar mund të
vrasin qelizën e infektuar dhe në këtë mënyrë e pengojnë përhapjen e
infeksionit. Limfocitet ndihmëse T janë të përfshira në formimin e
LTC-ve dhe në asistimin e limfociteve B për të prodhuar antitrupa.
Përveç kësaj, limfokinet e sekretuara nga limfocitet T mund të
rekrutojnë dhe aktivizojnë makrofagët dhe qelizat VN, në këtë mënyrë
duke e mobilizuar një sulm të përbashkët ndaj virusit.
|
| |
PËRMBLEDHJA E
MBROJTJEVE
Tabela 7 (Adaptuar nga: Baron, Medical
Microbiology, 2nd Ed., Tabela 69-2) përmbledh mbrojtjet e strehuesit
kundër infeksoneve virale dhe tregon caqet për të gjitha këto mbrojtje.
|
Tabela 7
Funksionet efektore të strehuesit në infeksionet virale
|
|
Mbrojtja e strehuesit |
Efektori |
Caku i efektorit |
|
Përgjigjet e hershme
jospecifike |
Ethet
Fagocitoza
Inflamacioni
Aktiviteti i qelizave VN
Interferoni |
Riprodhimi i virusit
Virusi
Replikimi i virusit
Qeliza e infektuar me virus
Riprodhimi i virusit, imunomodulimi |
|
Përgjigjet imune të
ndërmjetësuara nga qelizat |
Linfocitet T citotoksike
Makrofagët e aktivizuar
Limfokinet
ADCC |
Qeliza e infektuar me virus
Virusi, qeliza e infektuar me virus
Qeliza e infektuar me virus, imunomodulimi
Qeliza e infektuar me virus |
|
Përgjigjet imune humorale |
Antitrupi
Antitrupi + komplementi |
Virusi, qeliza e infektuar me
virus
Virusi, qeliza e infektuar me virus |
KONTRIBUTET
RELATIVE TË MEKANIZMAVE MBROJTËS TË STREHUESIT
Kontributet relative të mekanizmave të ndryshëm mbrojtës
të strehuesit do të varen nga natyra e virusit dhe porta e hyrjes.
Antitrupat do të jenë më të rëndësishëm në infeksionet në të cilat
viremia është karakteristika më e theksuar. Megjithatë, antitrupat mund
të mos jenë të dobishëm në infeksionet me herpes ose paramyxovirus, në
të cilat virusi mund të kalojë nga qeliza në qelizë përmes fuzionit të
qelizave. Në këtë rast, imuniteti i ndërmjetësuar nga qelizat është më i
rëndësishëm. Nëse virusi i infekton qelizat vetëm në sipërfaqen e
mukozave, atëherë rëndësi kanë antitrupat IgA.
Të kuptuarit e mekanizmave mbrojtës të strehuesit është i rëndësishëm në
zhvillimin e vaksinave dhe për administrimin e duhur të vaksinave. Nëse
antitrupat IgA janë të rëndësishëm për mbrojtje kundër virusit të
caktuar, atëherë vaksina duhet të jetë në gjendje të stimulojë prodhimin
e antitrupave IgA në sipërfaqen e duhur mukozave. Në mënyrë alternative,
nëse LTC-të janë të rëndësishme, atëherë vaksina duhet të jetë në
gjendje që të stimulojë prodhimin e LTC. Kjo është arsyeja pse vaksinat
e gjalla janë shpesh të preferuara kundrejt vaksinave te vdekura, sepse
vaksinat e gjalla zakonisht çojnë në gjenerimin e LTC-ve përderisa
vaksinat e vdekura nuk e bëjnë një gjë të tillë.
|
| |
IMUNOPATOLOGJIA E
NXITUR NGA VIRUSI
Ndonëse strehuesi ka një mori të mbrojtjeve për t’i përballuar
infeksioneve virale, nganjëherë shkaku kryesor i dëmtimit të indit është
pikërisht përgjigja imunologjike ndaj infeksionit. P.sh. infantët e
infektuar me citomegalovirus kanë imunokomplekse në qarkullim të gjakut,
të cilat depozitohen në veshka dhe nyje duke rezultuar në artrit dhe
glomerulonefrit. Shembull tjetër është sindromi i shokut hemorragjik
fatal, që ndërlidhet me ethet tropikale. Në këtë rast fiksimi i
komplementit nga ana e imunokomplekseve qarkulluese rezulton me lirimin
e produkteve të kaskadës së komplementit që çon në rritjen e befasishme
të lëshueshmërisë së enëve të gjakut, shok dhe vdekje.
IMUNOSUPRESIONI
Shumë virusë janë të aftë që të pengojnë përgjigjen imune dhe kështu të
tejkalojnë ose minimizojnë mbrojtjen e strehuesit. Shembulli më i mirë
është HIV, i cili infekton qelizat CD4+ dhe në këtë mënyrë shkatërron
sistemin imun specifik. Virusët tjerë (p.sh. virusi i morbillit) mund
gjithashtu të infektojnë limfocitet dhe të ndikojnë në riprodhimin dhe
diferencimin e tyre. Disa nga mekanizmat përmes të cilëve virusët mund
të shmangin mbrojtjen e strehuesit janë dhënë në tabelën 8 (Adaptuar nga:
Roitt, Immunology 5th Ed., Fig 16.10).
|
Tabela 8
Prodhimet virale që interferojnë me mbrojtjet e strehuesit |
|
Mbrojtja e prekur e
strehuesit |
Virusi |
Prodhimi i virusit |
Mekanizmi |
|
Interferoni |
EBV |
EBERS (ARN-të
e vogla) |
Bllokon aktivizmin e protein-kinazës |
|
Vaccinia |
eIF-2alfa
homolog |
Pengon fosforilimin e
eIF-2alfa përmes proteinë- kinazës |
|
Komplementi |
Vaccinia |
Homologe e
proteinave kontrolluese të komplementit |
Bllokon aktivizimin e
komplementit |
|
Antitrupi |
HSV-1 |
gE/gI |
Lidh
Fc-gama dhe bllokon funksionin |
|
Citokinet |
Myxoma |
Homolog me
receptorin e
IFN-gama |
Konkurron me IFN-gama dhe
bllokon funksionin |
|
Shope fibroma virus |
FNT
receptor |
Konkurron me FNT dhe bllokon
funksionin |
|
EBV |
IL-10
homolog |
Zvogëlon funksionin e
IFN-gama |
|
MHC Klasa I |
CMV |
Proteina e
hershme |
Pengon transportin e MHC të
ngarkuar me peptide |
|
Adenovirus |
E3 |
Bllokon
transportin e MHC për në sipërfaqe |
|
Apoptoza |
Adenovirus |
14.7K |
Inhibon kapsazat |
|
EBV |
Bcl-2 homolog |
Anti-apoptoik |
|
Qelizat VN |
HCMV |
UL-18 |
MHC homolog
inhibon qelizat VN |
|
|
|
 Kthehu
në seksionin e virologji të Microbiology and Immunology On-line
Kthehu
në seksionin e virologji të Microbiology and Immunology On-line
faqja së fundi e ndërruar në
Monday, February 16, 2015
faqja mbahet nga
Richard Hunt
|
 Figura
1 Zbulimi i interferonit
Figura
1 Zbulimi i interferonit Figura 3
Figura 3 Figura
4
Figura
4 Figura
5
Figura
5