|
xx |
x |
|
 |
 |
|
DOENÇAS
INFECCIOSAS |
BACTERIOLOGIA |
IMUNOLOGIA |
MICOLOGIA |
PARASITOLOGIA |
VIROLOGIA |
|
|
VIROLOGIA – CAPÍTULO CINCO
GENÉTICA VIRAL
Dr Margaret Hunt
Professor Emerita
Department of Pathology, Microbiology and Immunology
University of South Carolina School of Medicine
Tradução:
PhD. Myres Hopkins
|
|
En
Español |
|
|
|
NË SHQIPTARE |
Dê a
sua opinião
CONTATO |
|
BUSCA |
|
|

 |
|
|
|
|
|
OBJETIVOS
Introdução à genética de vírus animais
|
GENERALIDADES
Vírus crescem rapidamente,
há normalmente um grande número de virions na progênie por célula. Há,
entretanto mais chances de mutações ocorrendo em um curto período de tempo.
A natureza do genoma
viral (RNA ou DNA; segmentado ou não segmentado) tem papel importante na
genética do vírus.
Vírus podem mudar
geneticamente devido a mutações ou
recombinação
MUTANTES
Origem
Mutações espontâneas
Estas aparecem
naturalmente durante a replicação viral: ex. devido a erros da polimerase
replicadora do genoma ou como resultado da incorporação de formas
tautoméricas das bases
Vírus de DNA tendem a
ser mais estáveis geneticamente do que vírus de RNA. Há mecanismos de correção
de erro na célula hospedeira para reparo de DNA, mas provavelmente não para RNA.
Alguns vírus de RNA
são bastante invariantes por natureza. Provavelmente esses vírus têm a mesma
taxa elevada de mutação que outros vírus de RNA, mas são tão precisamente
adaptados para a transmissão e replicação que mudanças muitissimamente pequenas
resultam em sua falha em competir com sucesso com o vírus tipo parental (tipo
selvagem).
Mutações que são
induzidas por agentes físicos ou químicos compreendem
Químicos:
Agentes que agem
diretamente nas bases, ex. ácido nitroso
Agentes que agem indiretamente, ex. análogos de base que provocam êrros de
pareamento mais frequentemente do que as bases normais, gerando assim mutações.
Físicos:
Agentes tais como luz UV
ou raios-X
Tipos de
mutação
Mutantes podem ter
mutações de ponto (uma base substituida por outra) ou mutações por
inserção/deleção.
Exemplos
de tipos de mudanças fenotípicas vistas em vírus mutantes
(fenótipo = propriedades
observadas de um organismo)
Mutantes
condicionais letais:
Esses mutantes se
multiplicam em certas condições mas não em outras (enquanto que o vírus do tipo
selvagem cresce em ambas as condições)
ex. mutantes sensíveis à temperatura (ts) – Estes irão crescer em baixa
temperatura ex. 31 graus C mas não em ex. 39 graus C, tipo selvagem
cresce a 31 e 39 graus C. Aparentemente a razão para isso é frequentemente que a
proteína alterada não consegue manter a conformação funcional na temperatura
elevada.
ex. Mutação de
espectro de hospedeiro – Esses mutantes irão crescer somente em um grupo
de tipos celulares no qual o vírus do tipo selvagem irá crescer – tais
mutantes proporcionam um meio de investigar o papel da célula hospedeira na
infecção viral
Tamanho da placa:
As placas
podem ser maiores ou menores do que o vírus do tipo selvagem, às vezes
tais mutantes exibem patogenicidade alterada
Resistência a
droga:
Isto é importante no
desenvolvimento de agentes antivirais – a possibilidade de mutantes resistentes
aparecerem sempre deve ser considerada
Mutantes
deficientes em enzimas:
Algumas enzimas virais nem
sempre são essenciais e podemos isolar mutantes viáveis deficientes em enzimas;
ex. a
timidina cinase do vírus do herpes simplex normalmente não é necessária em
cultura de tecidos mas é importante na infecção de células neuronais
Mutantes
"Quentes":
Estes crescem melhor em temperaturas elevadas do que o vírus do
tipo selvagem.
Podem ser mais virulentos pois a febre do hospedeiro tem pouco efeito nos
mutantes mas tornam a replicação dos vírions do tipo selvagem mais lenta.
Mutantes atenuados:
Muitos mutantes virais provocam sintomas muito mais amenos (ou ausência de
sintomas) comparados com o vírus parental – a isso se diz que eles são
atenuados. Eles têm um potencial papel no desenvolvimento de vacinas e são
ferramentas úteis na determinação do por que o vírus parental é prejudicial.
|
 Figura 1 Recombinação por escolha de cópia
Figura 1 Recombinação por escolha de cópia |
TROCA DE
MATERIAL GENÉTICO
Recombinação
Troca de informação
genética entre dois genomas.
Recombinação
"Clássica"
Esta envolve a quebra de
ligações covalentes dentro do ácido nuclêico, troca de informação genética, e
re-formação das ligações covalentes.
Este tipo de
quebra/reunião é comum em vírus de DNA ou naqueles vírus de RNA que têm uma fase
de DNA (retrovírus). A célula hospedeira tem sistemas de recombinação de DNA.
Recombinação deste tipo e muito rara em vírus de RNA (provavelmente não há
enzimas do hospedeiro para recombinação de RNA). Picornavívus exibe uma forma de
recombinação de muito baixa eficiência. O mecanismo não é idêntico ao mecanismo
padrão para DNA, e é provavelmente um mecanismo do tipo "escolha
de cópia" (figura 1) no qual a polimerase troca de templates quando copia o
RNA.
|
 Figura
2 Resgate de marcador Figura
2 Resgate de marcador |
A recombinação é
também comum nos coronavírus – e também o mecanismo é diferente do que
ocorre com DNA e provavelmente é uma consequência da maneira não usual como
o RNA é sintetizado neste vírus.
Até o presente, não
há evidência de recombinação nos vírus de RNA de fita negativa produzindo
vírus viáveis (Nesses vírus, o RNA genômico é empacotado em nucleocapsídeos
e não está prontamente disponível para pareamento de bases).
Vários usos de
técnicas de recombinação
a) Mapeamento de
genomas (quanto mais separados um do outro forem dois genes, maior a chance
de eventos recombinação entre eles).
b) Resgate de
marcador – Fragmentos de DNA de vírus do tipo selvagem podem
recombinar com vírus mutantes para gerar vírus do tipo selvagem -
isso proporciona um meio de conferir uma função gênica a uma região
específica do genoma. Este fato também proporciona um meio de inserir
material exógeno em um gene (figura 2).
Recombinação habilita um vírus a pegar informação genética de vírus do mesmo
tipo, e ocasionalmente de vírus não relacionados ou mesmo do genoma
hospedeiro (como ocorre em alguns retrovírus – ver
retrovírus).
|
 Figura 3 Ressortimento de genoma viral em vírus segmentados
Figura 3 Ressortimento de genoma viral em vírus segmentados |
Ressortimento (sentido:
re-embaralhamento)
Se um vírus tem um genoma
segmentado e se duas variantes daquele vírus infecta uma única célula, a
progênie de virions pode resultar de alguns segmentos de um parental, e outros
do outro.
Este é um processo
eficiente – mas é limitado a vírus com genomas segmentados – até o presente os
únicos vírus humanos caracterizados com genomas segmentados são os vírus de RNA
ex. ortomixovírus, reovírus, arenavírus, buniavírus.
Ressortimento pode ter
um papel importante na natureza na geração de novas combinações e também tem
sido útil em experimentos laboratoriais (figura 3). Tem sido também explorada a
atribuição de funções a diferentes segmentos do genoma. Por exemplo, em um vírus
ressortido, se um segmento vem do vírus A e o resto do vírus B, podemos ver
quais as propriedades que se assemelham à do vírus A e quais as do vírus B.
Ressortimento é um tipo de recombinação não clássica.
|
 Figura 4 Ressortimento de genes entre uma linhagem atenuada do vírus da
influenza e uma linhagem nova virulenta na formação de uma vacina
atenuada contra a influenza (link
para o capítulo de vacina em Inglês)
Adaptado de: Treanor JJ Infect.
Med. 15:714
Figura 4 Ressortimento de genes entre uma linhagem atenuada do vírus da
influenza e uma linhagem nova virulenta na formação de uma vacina
atenuada contra a influenza (link
para o capítulo de vacina em Inglês)
Adaptado de: Treanor JJ Infect.
Med. 15:714 |
Genética aplicada
Existe uma vacina chamada Flumist (LAIV, aprovada em Junho de
2003) para o vírus da influenza que envolve alguns dos princípios discutidos
acima. A vacina é trivalente – ela contém 3 linhagens de vírus da influenza:
Os vírus são
de linhagens adaptadas ao frio que podem crescer bem a 25 graus C e porisso
podem crescer no trato respiratório superior onde é mais frio. Os vírus são
sensíveis a temperatura e crescem pouco no trato respiratório inferior e mais
aquecido. Os vírus são de linhagens atenuadas e muito menos patogênicas do que o
vírus do tipo selvagem. Isso se deve a várias mudanças nos vários segmentos do
genoma.
Anticorpos
para proteínas de superfície do vírus da influenza (HA -
hemaglutinina e NA -
neuraminidase) são importantes na proteção contra
infecção. A HA e a NA mudam a cada ano. A tecnologia da vacina usa ressortimento
para gerar vírus ressortidos que têm seis segmentos gênicos do vírus atenuado
adaptado ao frio e os segmentos codificadores HA e NA do vírus, o que tem chance
de ser um problema na estação da influenza que está por vir.
Esta vacina é
uma vacina viva e é dada intranasalmente como um spray e pode induzir a
imunidade mucosa e sistêmica.
|
| |
Complementação
Interação a um nível
funcional NÃO ao nível do ácido nuclêico. Por exemplo, se pegarmos dois mutantes
com uma lesão ts (sensível à temperatura) em diferentes genes, nenhum
pode crescer a uma temperatura elevada (não permissiva). Se infectarmos a mesma
célula com ambos os mutantes, cada mutante pode fornecer a função que falta do
outro e portanto eles podem replicar (entretanto, a progênie de virions irá
ainda conter genomas mutantes ts e irão ser sensíveis à temperatura).
Podemos usar
complementação para agrupar mutantes ts, pois mutantes no mesmo gene não
serão capazes normalmente de complementar um ao outro. Esta é uma ferramenta
básica em genética para determinar se mutações estão no mesmo ou em genes
diferentes e para determinar o número mínimo de genes que afetam uma função.
|
| |
Multiplicidade de
reativação
Se
vírus de DNA dupla fita forem inativados usando irradiação ultravioleta, nós
frequentemente vemos reativação se infectarmos células com o vírus inativado a
uma multiplicidade de infecção muito alta (ou seja, várias partículas virais por
célula) – isso ocorre porque vírus inativados cooperam de alguma forma.
Provavelmente complementação permite que vírus cresçam inicialmente, pois genes
inativados em um virion podem ainda estar ativos em um dos outros. Como o número
de genomas presentes aumenta devido a replicação, a recombinação pode ocorrer,
resultando em novos genótipos, e às fezes regenerando o vírus do tipo
selvagem.
Vírus defeituosos
Vírus defeituosos não
têm o complemento inteiro de genes necessários para um ciclo de infecção
completa (muitos são mutantes por deleção) – e assim eles necessitam de outro
vírus para fornecer as funções que faltam – este segundo vírus é chamado de
vírus auxiliar.
Vírus defeituosos
fornecem os sinais necessários para a polimerase replicar seus genomas e para
seus genomas ser empacotados, e mais nada. Alguns vírus defeituosos conseguem
fazer mais coisas sozinhos.
Alguns exemplos de
vírus defeituosos:
Some
retroviruses have picked up host cell sequences but have lost some viral
functions. These need a closely related virus which retains these functions as a
helper.
Alguns vírus defeituoses
podem usar vírus não relacionados como auxiliares. Por exemplo, o
vírus da hepatitis delta (um vírus de RNA)
não codifica para suas próprias proteínas do envelope mas usa o envelope do
vírus da hepatite B (um vírus de DNA).
Partículas
defeituosas interferentes
A replicação dos vírus
auxiliares pode ser menos eficiente do que se o vírus (partícula) defeituoso não
estivesse lá. Isso é porque a partícula defeituosa está competindo com a
auxiliar pelas funções que a auxiliar proporciona. Este fenômeno é conhecido
como interferência, e partículas defeituosas que provocam este fenômeno são
conhecidas como partículas “defeituosas interferentes” (DI). Nem todos os vírus
defeituosos fazem isso, mas muitos fazem.
Note que é possível que partículas defeituosas interferentes podem modular
infecções naturais
|
 Figura 5 Mistura fenotípica entre dois vírus diferentes que infectam a
mesma célula
Figura 5 Mistura fenotípica entre dois vírus diferentes que infectam a
mesma célula |
Mixagem fenotípica
Se dois vírus diferentes
infectam uma célula, os vírus da progênie podem conter componentes da capa
derivados de ambos os parentais, e assim eles irão ter propriedades de capa de
ambos os parentais.
Isso é chamado
de mixagem fenotípica (figura 5).
ISSO NÃO ENVOLVE ALTERAÇÃO
NO MATERIAL GENÉTICO, a progênie de tais virions será determinada pelo tipo de
genoma parental que é empacotado e não pela natureza do envelope.
Mixagem fenotípica pode ocorrer entre vírus relacionados, ex. diferentes membros
da família dos Picornavírus, ou entre vírus geneticamente não relacionados, ex.
vírus Rabdo e Paramixo. No último caso os dois vírus envolvidos são normalmente
envelopados pois ao que parece há menor controle no empacotamento de
nucleocapsídeos em outros envelopes virais do que no empacotamento de ácidos
nuclêicos em outros capsídeos icosaédricos de outros vírus.
|
 Figura 6 Mistura fenotípica para formar um pseudotipo
Figura 6 Mistura fenotípica para formar um pseudotipo |
Podemos ter também a
situação em que uma capa é inteiramente a mesma de outro vírus, ex. um
nucleocapsídeo de retrovírus em um envelope de rabdovírus. Esse tipo de mixagem
fenotípica é às vezes referida como uma formação de pseudotipo (pseudovirion)
(figura 6). O pseudotipo descrito acima irá mostrar as características de
antigenicidade em termos de adsorção-penetração-superfície do rabdovírus, e irá
então com a infecção se comportar como um retrovírus e produzir uma progênie de
retrovírus. Isso resulta em pseudotipo, tendo um tropismo espectro/tecido
alterado em base temporária.
|
|
|
 Voltar
à Seção Virologia do Microbiologia e Imunologia Online
Voltar
à Seção Virologia do Microbiologia e Imunologia Online
This page last changed on
Monday, February 08, 2021
Página mantida por
Richard Hunt
|
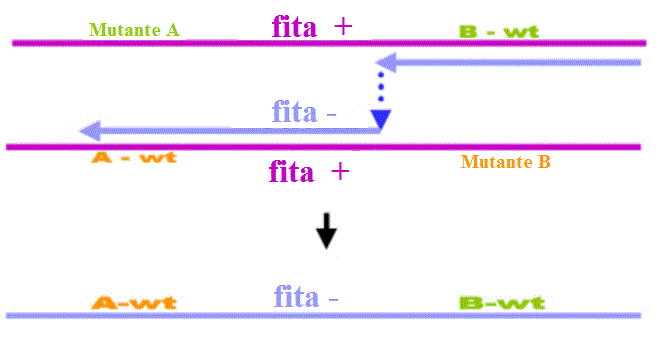 Figura 1 Recombinação por escolha de cópia
Figura 1 Recombinação por escolha de cópia Figura
2 Resgate de marcador
Figura
2 Resgate de marcador 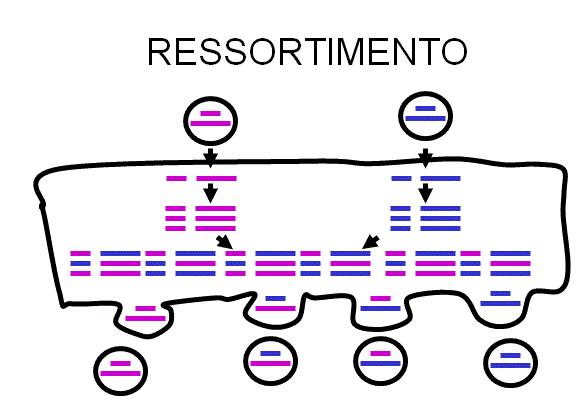 Figura 3 Ressortimento de genoma viral em vírus segmentados
Figura 3 Ressortimento de genoma viral em vírus segmentados Figura 4 Ressortimento de genes entre uma linhagem atenuada do vírus da
influenza e uma linhagem nova virulenta na formação de uma vacina
atenuada contra a influenza (link
para o capítulo de vacina em Inglês)
Adaptado de: Treanor JJ Infect.
Med. 15:714
Figura 4 Ressortimento de genes entre uma linhagem atenuada do vírus da
influenza e uma linhagem nova virulenta na formação de uma vacina
atenuada contra a influenza (link
para o capítulo de vacina em Inglês)
Adaptado de: Treanor JJ Infect.
Med. 15:714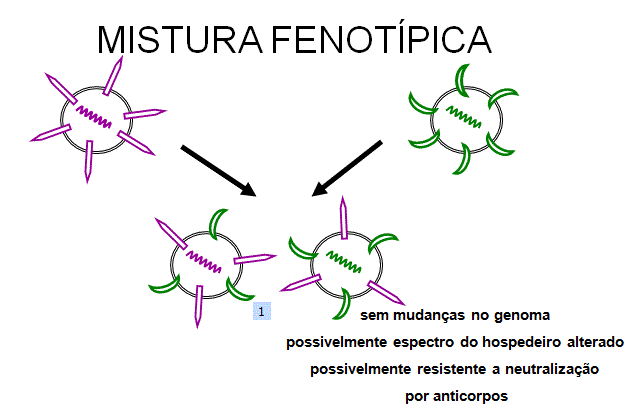 Figura 5 Mistura fenotípica entre dois vírus diferentes que infectam a
mesma célula
Figura 5 Mistura fenotípica entre dois vírus diferentes que infectam a
mesma célula Figura 6 Mistura fenotípica para formar um pseudotipo
Figura 6 Mistura fenotípica para formar um pseudotipo