|
x |
x |
|
 |
 |
|
x |
BACTERIOLOGÍA |
INMUNOLOGÍA
|
MICOLOGÍA |
PARASITOLOGÍA |
VIROLOGÍA |
|
EN INGLÉS |
BACTERIOLOGÍA - CAPÍTULO OCHO
INTERCAMBIO DE INFORMACIÓN GENÉTICA
Dr Gene
Mayer
Professor Emeritus
University of South Carolina School of Medicine
Traducido por :
Dr. en C. Paula Figueroa-Arredondo
|
|
VA EL CAPÍTULO 9 |
|
|
|
INSTITUTO POLITÉCNICO NACIONAL |
E-MAIL
DR PAULA FIGUEROA |
|
OBJETIVOS DE APRENDIZAJE
Explicar los mecanismos de la transferencia de genes en bacterias.
Describir la naturaleza de los elementos genéticos transponibles y los
plásmidos.
Discutir la importancia de la transferencia genética, los elementos
genéticos transponibles y los plásmidos |
INTRODUCCIÓN
En
las poblaciones bacterianas constantemente están surgiendo mutaciones a
causa de los errores que aparecen durante la replicación. Si existe
cualquier ventaja selectiva para una mutación en particular (ej.
resistencia a antibióticos), la mutante enseguida se convertirá en el
principal componente de la población, debido a la rápida tasa de crecimiento
de las bacterias. Además, dado que las bacterias son organismos haploides,
aún las mutaciones que normalmente podrían ser recesivas serán expresadas.
Por ello, en poblaciones bacterianas las mutaciones pueden representar un
problema en el tratamiento de las infecciones causadas por bacterias. No
solo son problema las mutaciones, las bacterias tienen mecanismos por los
cuales sus genes pueden transferirse de una célula a otra. Por tanto, una
mutación que surge en una célula siempre puede ser transmitida a las otras.
La
transferencia genética en bacterias es unidireccional y va de una célula
donadora a una receptora y la donadora usualmente cede solamente una pequeña
parte de su DNA a la receptora. Por tanto, no se forman cigotos completos;
en su lugar se forman cigotos parciales (merocigotos).
Los
genes bacterianos usualmente se transfieren a miembros de la misma especie
aunque ocasionalmente puede ocurrir transferencia hacia otras especies. La
Figura 1 ilustra las transferencias genéticas que se ha demostrado que
ocurren entre especies bacterianas diferentes.
|
PALABRAS CLAVE
Merocigoto
Transformación
Competencia
Recombinación Homóloga
Transducción
Transducción generalizada
Transducción especializada
Conversión lisogénica
Conjugación
Pilus F/pili sexual
Replicón
F+
F-
Hfr
F'
Elemento genético transponible
Secuencias de inserción
Transposón
Recombinación sitio-específico
Variación de fase
Plásmidos, Plásmido conjugativo
Plásmido No conjugativo
Factor R
RTF
Determinante R |
MECANISMOS DE TRANSFERENCIA GENÉTICA EN BACTERIAS
Transformación
La
transformación es la transferencia genética que resulta de la incorporación
de DNA desnudo por una célula receptora desde una célula donadora.
Ciertas bacterias (ej.
Bacillus, Haemophilus, Neisseria, Pneumococcus)
son capaces de tomar DNA del medio ambiente y ese DNA que es introducido
puede llegar a ser incorporado al cromosoma de la célula bacteriana
receptora.
Factores que afectan la transformación
a.
Tamaño del DNA
Funciona mejor el DNA de doble cadena de al menos 5
X 105 daltones. Por tanto, la transformación es sensible a las
nucleasas del medio ambiente.
b.
Competencia de la célula receptora
Algunas bacterias son capaces de
incorporar DNA en forma natural. Sin embargo, estas bacterias solo toman al
DNA en una etapa particular de su ciclo celular, cuando producen una
proteína específica llamada factor de competencia. Cuando la bacteria
se encuentra en este estadio se dice que es competente.
Otras
bacterias no son capaces de incorporar el DNA naturalmente, sin embargo en
estas bacterias la competencia puede ser inducida in
vitro
mediante tratamiento con sustancias químicas
(ej.
CaCl2).
Pasos de la transformación.
a.
Incorporación del DNA
La incorporación del DNA por las bacterias
Gram+ y Gram- es diferente. En las bacterias Gram+ el DNA se introduce en
forma de moléculas de cadena sencilla y la cadena complementaria se
sintetiza dentro de la célula receptora. En contraste, las bacterias Gram-
incorporan DNA de doble cadena.
b.
Recombinación General/Legítima/Homóloga
Luego de que el DNA de la célula donadora se ha incorporado, ocurre un evento de
recombinación recíproca entre el cromosoma y el DNA de la célula donadora.
Esta recombinación requiere de que exista homología entre el DNA del donador
y el cromosoma receptor, lo que finalmente resulta en la substitución de DNA
entre la receptora y la donadora, como se ilustra en la Figura 2.
|
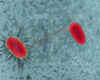 E. coli
(bacilo procariote) cepas llevandoa cabo la conjugación.
Una cepa tiene fimbrias
©
Dr
Dennis Kunkel,
University of Hawaii. Used with permission
E. coli
(bacilo procariote) cepas llevandoa cabo la conjugación.
Una cepa tiene fimbrias
©
Dr
Dennis Kunkel,
University of Hawaii. Used with permission
 Figura 1 Transferencia genética que se ha demostrado que ocurre entre
diferentes especies de bacterias.
Figura 1 Transferencia genética que se ha demostrado que ocurre entre
diferentes especies de bacterias.
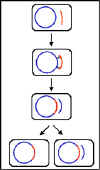 Figura 2 Recombinación General. El DNA del donador se muestra en rojo y
el del receptor en azul.
Figura 2 Recombinación General. El DNA del donador se muestra en rojo y
el del receptor en azul.
 Figura 3 El mecanismo de la transducción generalizada.
Figura 3 El mecanismo de la transducción generalizada. |
Esta recombinación requiere de los genes de la recombinación bacteriana
(recA, B y C) y de que exista homología entre los DNAs involucrados.
Este tipo de recombinación se denomina recombinación general,
legítima u homóloga. Debido al requerimiento de homología entre las
células donadora y huésped, solo el DNA de una bacteria cercanamente
relacionada se esperaría que transformara exitosamente, aunque en raras
ocasiones se ha demostrado que sí ocurre transferencia genética de este
tipo entre bacterias relacionadas de forma más bien distante.
Importancia
La transformación ocurre en la naturaleza de
manera normal y es un mecanismo que puede conducir al incremento de la
virulencia bacteriana. Por otra parte, la transformación in vitro
ha sido ampliamente utilizada en la tecnología del DNA recombinante.
Transducción
La transducción es la transferencia de información genética desde un
donador a un receptor y está mediada por un bacteriófago (fago).
La cubierta del fago protege al DNA del medio ambiente, así es que la
transducción, a diferencia de la transformación, no se ve afectada por
las nucleasas en el medio ambiente. No todos los fagos pueden mediar la
transducción. En la mayoría de los casos la transferencia genética se
realiza entre miembros de las mismas especies bacterianas. Sin embargo,
si un fago en particular posee un amplio rango de huéspedes que él es
capaz de infectar, entonces la transferencia entre las especies puede
ocurrir. La capacidad del fago para mediar la transducción, está
relacionada con el ciclo de vida del mismo.
Tipos de
Transducción
a. Transducción Generalizada
La transducción generalizada es el
mecanismo por el cual potencialmente cualquier gene bacteriano de la
donadora puede ser transferido a la célula receptora. El mecanismo de la
transducción generalizada se ilustra en la Figura 3.
Los fagos que median la transducción generalizada, normalmente cortan el
DNA de la célula huésped en pequeñas piezas y empacan ambos DNAs al
interior de la partícula fágica mediante un mecanismo llamado “head
full” o llenado de las cabezas del fago. Ocasionalmente una de las
piezas del DNA de la bacteria huésped resulta empacada al azar dentro de
una cubierta de fago. Por lo tanto cualquier gene de la bacteria
donadora puede ser potencialmente transferido, pero solamente se
transferirá tanto DNA como pueda caber en una sola cápside. Cuando la
célula receptora se infecta con un fago que contiene DNA de una
donadora, el DNA de la donadora puede entrar a la receptora. Ya dentro
de la célula receptora puede ocurrir el evento de la recombinación
generalizada, en el cual se substituye el DNA de la célula donadora por
el de la receptora (Figura 2).
|
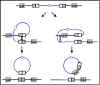 Figura 4. El mecanismo de la transducción especializada.
Figura 4. El mecanismo de la transducción especializada. |
b. Transducción especializada
La transducción especializada es la
transducción en la cual solo ciertos genes del donador pueden ser
transferidos al receptor. Diferentes fagos pueden transferir diferentes
genes pero un fago individual solamente puede transferir unos pocos
genes. La transducción especializada está mediada por fagos lisogénicos
o fagos temperados y los genes que se llegan a transferir dependerán del
lugar donde el profago queda insertado en el cromosoma. El mecanismo de
la transducción especializada se ilustra en la Figura 4.
Durante la
escisión (separación) del profago, un error llega a
ocurrir ocasionalmente en el cual un poco del DNA del huésped escinde
(se separa del cromosoma) junto con el DNA del fago. Solo puede ser
transferido el DNA del huésped que esté flanqueando cada lado del sitio
donde el profago se ha insertado, (ej. transducción
especializada). Después de la replicación y la liberación del fago y a
través de la infección de la célula receptora, puede ocurrir una lisogenización de la receptora dando como resultado la transferencia
estable de los genes de la donadora. La receptora ahora tendrá dos
copias de los genes que le fueron transferidos. También es posible que
se lleve a cabo una recombinación legítima entre los genes de la
donadora y de la receptora.
Importancia
La conversión lisogénica (mediada por fago) ocurre en
la naturaleza y es la fuente de donde proceden las cepas virulentas.
|
|
PELÍCULA
Conjugación
Alta resolución
Baja resolución
© Mondo Media, San Francisco, Calif., USA and and
The MicrobeLibrary
Este video clip demuestra el proceso de la conjugación. Primero,
dos bacterias recombinan su material vía el pilus sexual. Luego, una
cadena del plásmido se transfiere a la célula unida. Note que el
plásmido original no se pierde en la primera célula. Finalmente, cada
célula duplica inmediatamente la cadena sencilla de modo que ambas
bacterias tienen una copia del plásmido de cadena doble.
|
Conjugación
La conjugación es la transferencia de DNA de una donadora a una
receptora, mediante contacto físico directo entre las células. En las
bacterias existen dos tipos de células y son las donadoras (macho) y las
receptoras (hembras) y la dirección de la transferencia genética es en
un solo sentido; así el DNA se transfiere desde la donadora hacia la
receptora.
Tipos de células acopladas (mating cells) en las bacterias
a.
Donadora
La capacidad de una bacteria de ser el donador es
consecuencia de la presencia en dicha célula de una pieza extra de DNA,
llamada factor F, factor de fertilidad o factor sexual. El
factor F es una pieza circular de DNA que es capaz replicar en forma
autónoma en la célula; es un replicón independiente. Las piezas de DNA
extra-cromosomal que pueden replicar autónomamente, reciben el nombre
genérico de plásmidos. El factor F posee los genes necesarios
tanto para su replicación como para su habilidad de transferir DNA a la
célula receptora. Una de las cosas que el factor F codifica es la
capacidad de producir una estructura llamada pilus sexual (pilus F) sobre la superficie de la bacteria.
Este pilus es
importante en el proceso de conjugación.
El factor F no es el único plásmido que puede mediar la conjugación
pero generalmente se toma como modelo.
b.
Receptora
La capacidad de actuar como receptora es
consecuencia de la carencia de esta célula del factor F.
|
| |
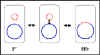 a
a
 b
b
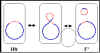
Figura 5 Estados fisiológicos del factor F.r
|
Estados fisiológicos del factor F
a.
(F+) Autónomo
El factor F en este estado
lleva solamente aquellos genes necesarios para su replicación y para la
transferencia de DNA. No hay genes cromosomales asociados con el factor
F en las cepas F+.
En los cruces del tipo F+ X F- el F- se
convierte en F+ mientras que F+ permanece como F+.
Por lo tanto, el factor F es infeccioso. Por otra parte, solamente se
presenta un bajo nivel de transferencia de los genes cromosomales de la
donadora.
b.
(Hfr) Integrado
En este estado el factor F se
encuentra integrado en el cromosoma bacteriano, vía un evento de
recombinación, como se ilustra en la Figura 5a
En los cruces del tipo Hfr X F- el F- raramente se
convierte en Hfr y la célula Hfr permanece como tal. En éste caso existe
una alta frecuencia de transferencia de los genes cromosomales del
donador, de ahí que el nombre de la cepa sea hfr, del inglés high frequency of recombination.
|
| |
c.
Autónomo con genes cromosomales (F')
En este estadio el
factor F es autónomo, pero ahora contiene algunos genes cromosomales.
Los factores F' se producen por escisión del factor F de una Hfr, como
se ilustra en la Figura 5b. Ocasionalmente, cuando el factor F se
escinde del cromosoma Hfr, los genes del donador localizados en cada
lado del factor F pueden escindir junto con el factor F generando una
F'. Los factores F' se denominan dependiendo de los genes cromosomales
que contienen.
En los cruces del tipo F' X F- el F- se convierte
a F' mientras que F' permance como tal. Por otra parte esta bacteria
presenta una alta frecuencia de transferencia de aquellos genes
cromosomales se encuentran en el F' y presenta una baja frecuencia de
transferencia de otros genes cromosomales del donador.
|
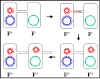 Figura 6 Mecanismo de los cruces F+ x F-
Figura 6 Mecanismo de los cruces F+ x F- |
Mecanismo de la conjugación
a. Cruces
F+ X F- (Figura 6)
i) Formación del par
La punta del pilus sexual se pone en contacto con
la receptora y formandose un puente de conjugación entre las
dos células. Es a través de este puente que el DNA pasará del
donador al receptor. De tal forma que el DNA queda protegido de las
nucleasas ambientales. Los pares de acoplamiento o mating pairs
pueden ser separados por fuerzas tan simples como la agitación y así la
conjugación se puede interrumpir. Consecuentemente, los pares de
apareamiento permanecen asociados solamente por un tiempo corto.
ii) Transferencia del DNA
El DNA del plásmido se corta en un sitio
específico llamado origen de la transferencia y se replica
mediante un mecanismo de círculo rodante. Una sola cadena de DNA pasa a
través del puente de conjugación y entra a la receptora donde la segunda
cadena se replica.
iii) Este proceso explica los cruces característicos F+ X F-.
La receptora se convierte en F+, la donadora permanece como F+
y presenta una baja frecuencia de transferencia de los genes
cromosomales del donador. Como se describe en la Figura 7, realmente no
hay transferencia de los genes cromosomales del donador. Sin embargo en
la práctica, existe un bajo nivel de transferencia de los genes
cromosomales del donador en tales cruces.
|
|
ANIMACIÓN
Apareamiento de
cepas Bacterianas F+ y F-
© Thomas M. Terry, University of Connecticut, Storrs, Conn., USA and
The
MicrobeLibrary
El plásmido F es un plásmido auto-transmisible que se encuentra en
algunas cepas de E. coli. Las células que poseen uno o mas copias
del plásmido F se denominan F+; las células carentes de plásmido F se
llaman F-. La animación ilustra varios estadios de la transferencia del
plásmido F desde las células F+ hacia las F-. |
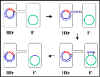 Figura 7 Mecanismo de los cruces Hfr x F-
Figura 7 Mecanismo de los cruces Hfr x F- |
b. Cruces
Hfr X F- (Figura 7)
i) Formación del
Par
ii) Transferencia de DNA – El DNA sufre un corte en el sitio de origen
de la transferencia y se replica mediante un mecanismo de círculo
rodante. En este caso el DNA que se transfiere primero es el del
cromosoma. Dependiendo del lugar del cromosoma donde el factor F se ha
integrado y en qué orientación lo haga, diferentes genes cromosomales
serán transferidos a tiempos diferentes. Sin embargo, el orden relativo
y las distancias de los genes siempre permanecerán igual. Solo hasta
que el cromosoma entero se haya transferido, entonces el factor F se
transferirá. Ya que los movimientos tales como las fuerzas de agitación
son capaces de separar a los pares sexuales formados, es raro que el
cromosoma entero se transfiera. Por ello, la receptora generalmente no
recibe el factor F en un cruce Hfr X F-.
iii) Recombinación legítima – La recombinación entre el DNA transferido
y el cromosoma da como resultado un intercambio del material genético
entre la donadora y la receptora.
iv) Este mechanismo explica las caracteristicas de los cruces Hfr X F-.
La receptora permanece como F-, la donadora permanece Hfr y existe una
alta frecuencia de transferencia de los genes cromosomales a partir de
la donadora.
|
|
ANIMATION
Mating of Hfr and F- Bacterial Strains
© Thomas M. Terry, University of Connecticut, Storrs, Conn., USA
and
The
MicrobeLibrary
|
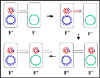 Figura 8 El mecanismo de cruces F" x F-
Figura 8 El mecanismo de cruces F" x F- |
c.
Cruces F' X F- (Figura 8)
i)
Formación del Par
ii)
Transferencia del DNA - Este proceso es similar al cruce F+ X F-.
Sin embargo, debido a que el F' lleva algunos genes cromosomales estos
también van a ser transferidos.
iii)
Recombinación homóloga. No es necesaria aunque puede ocurrir.
iv)
Este mecanismo explica las características de los cruces F' X F-.
El F- se convierte en F', el F' permanece como tal y la se presenta una alta
frecuencia de transferencia de los genes del donador que van en el F' pero
una baja frecuencia de transferencia de otros genes cromosomales del
donador.
4)
Importancia – Entre las bacterias Gram negativas esta es la forma principal
en que se transfieren los genes. La transferencia puede ocurrir entre
diferentes especies bacterianas. La transferencia de resistencia múltiple a
los antibióticos por conjugación ha llegado a ser un problema relevante en
el tratamiento de ciertas enfermedades bacterianas. Debido a que la célula
receptora se convierte en donadora después de la transferencia del plásmido,
es fácil ver por qué un gen de resistencia a los antibióticos que va en un
plásmido puede rápidamente convertir una población sensible de bacterias en
una resistente.
Las
bacterias Gram positivas también tienen plásmidos que llevan genes de
resistencia múltiple a los antibióticos, en algunos casos estos plásmidos se
transfieren por conjugación mientras que en otras ellos se transfieren por
transducción. El mecanismo de conjugación en las bacterias Gram + es
diferente al de las Gram -. En las bacterias Gram + el donador produce un
material adhesivo el cual causa agregación con el receptor y el DNA se
transfiere.
|
| |
ELEMENTOS GENÉTICOS TRANSPONIBLES
Elementos
Genéticos Transponibles
Los
elementos genéticos transponibles son segmentos de DNA que tienen la
capacidad de moverse desde una localización hasta otra (ej. genes
saltarines).
Propiedades de los Elementos Genéticos Transponibles
1.
Movimiento al azar- Los elementos genéticos transponibles pueden moverse
desde cualquier molécula de DNA a cualquier otra molécula de DNA o aún a
otro lugar dentro de la misma molécula. El movimiento no es totalmente al
azar; existen sitios preferentes en una molécula de DNA en la cual se
insertará el elemento genético transponible.
2.
Incapaz de auto-replicarse – Los elementos genéticos transponibles no
existen de manera autónoma (a excepción – algunos genes transponibles en los
fagos) y por tanto, para ser replicados deben ser parte de algún otro
replicón.
3.
Transposición mediada por recombinación sitio-específica – La transposición
requiere poca o ninguna homología entre la localización actual y el nuevo
sitio. El evento de transposición está mediado por una transposasa
codificada por el elemento genético transponible. La recombinación que no
requiere de homología entre las moléculas recombinantes se denomina de sitio-específico, ilegítima o bien recombinación no-homóloga.
4. Transposición acompañada por duplicación – En muchas instancias la
transposición del elemento genético transponible resulta en la remoción del
elemento de su sitio original y su inserción en un nuevo sitio. Sin embargo,
en algunos casos el evento de transposición se acompaña de una duplicación
del elemento genético transponible. Una copia permanece en el sitio original
y la otra se transpone en el nuevo sitio.
|
 Figura 9 Estructura de los elementos genéticos transponibles
Figura 9 Estructura de los elementos genéticos transponibles |
Tipos de Elementos Genéticos Transponibles
1.
Secuencias de Inserción (IS)
Las secuencias de inserción son elementos
genéticos transponibles que llevan genes desconocidos, con excepción de
aquellos que se requieren para la transposición.
a.
Nomenclatura
A las secuencias de inserción (insertion sequences)
se les da la designación IS seguida por un número (ej: IS1)
b. Estructura (Figura 9)
Las
secuencias de inserción son pequeños tramos de DNA que a sus extremos tienen
secuencias repetidas que están involucradas en la transposición. En medio de
las secuencias terminales repetidas hay genes involucrados en la
transposición y secuencias que pueden controlar la expresión de los genes,
pero no presentan otros genes que no sean esenciales.
c. Importancia
i)
Mutación – La introducción de una secuencia de inserción en medio de un gene
bacteriano resultará en la inactivación de tal gene.
ii)
Inserción de plásmidos en los cromosomas – Los sitios en los cuales los
plásmidos se insertan en el cromosoma bacteriano se localizan ya sea en las
propias secuencias de inserción o cerca de ellas.
iii)
Variación de Fase – Los antígenos flagelares son de los principales
antígenos ante los cuales se dirige la respuesta inmune, en nuestro intento
de luchar contra la infección bacteriana. En Salmonella existen genes
que codifican para dos genes flagelares antigenicamente diferentes. La
expresión de estos genes está regulada por secuencias de inserción. En una
orientación, uno de los genes está activo mientras que en la otra
orientación el otro gene flagelar estará activo. Como resultado Salmonella puede cambiar sus flagelos en respuesta al ataque del sistema
inmune. La variación de fase en los antígenos flagelares de Salmonella
no es única. También se ha visto que ocurre con otros antígenos de
superficie. Además el mecanismo de la variación de fase puede ser diferente
en diversas especies de bacterias (ej. transformación en Neisseria).
2. Transposones (Tn)
Los transposones son elementos genéticos
transponibles que llevan uno o más de otros genes además de aquellos que son
esenciales para la transposición.
|
 Figura 10 Estructura de un Transposón.
Figura 10 Estructura de un Transposón. |
a.
Nomenclatura – A los transposones se les da la designación
Tn seguida de un número.
b.
Estructura - La estructura de un transposón es similar a la de
una secuencia
de inserción. Los genes extra están localizados entre las secuencias
repetidas terminales. En algunas instancias (transposones compuestos)
las secuencias repetidas terminales son de hecho secuencias de
inserción. (See Figure 10).
c.
Importancia - Muchos genes de resistencia a los antibióticos
se localizan en transposones. Debido a que los transposones pueden
saltar de una molécula DNA a otra, estos transposones de resistencia a
antibióticos son un factor principal en el desarrollo de plásmidos los
cuales pueden conferir resistencia a múltiples drogas en las bacterias
que albergan tales plásmidos. Estos plásmidos de resistencia a múltiples
drogas han llegado a ser un problema médico grave, debido que el uso
indiscriminado de antibióticos ha dado lugar a una ventaja selectiva
presente en las bacterias que poseen este tipo de plásmidos.
|
| |
PLÁSMIDOS
Definición
Los plásmidos son elementos genéticos extra-cromosomales
que pueden llevar a cabo una replicación autónoma. Un episoma es un
plásmido que además es capaz de integrarse al cromosoma bacteriano.
Clasificación de los Plásmidos
1. Propiedades de Transferencia
a.
Plásmidos Conjugativos – Los plásmidos conjugativos son
aquellos que son los mediadores del proceso de la conjugación. Estos
plásmidos usualmente son grandes, tienen todos los genes necesarios para una
replicación autónoma y para la transferencia del DNA hacia una célula
receptora (ej. genes para el pilus sexual).
b.
Plásmidos No-conjugativos – Los plásmidos no-conjugativos son
aquellos que no pueden mediar el proceso de la conjugación. Estos son
plásmidos usualmente más pequeños que los plásmidos conjugativos y carecen
de uno o más de los genes necesarios para la transferencia del DNA. Un
plásmido no-conjugativo puede transferirse por conjugación si la célula
también alberga a un plásmido conjugativo.
2.
Efectos en el fenotipo
a.
Plásmido de Fertilidad (factor F)
b.
Plásmidos Bacteriocinogénicos - Estos plásmidos poseen genes
que codifican para substancias que matan a otras bacterias. Dichas
substancias se llaman bacteriocinas o colicinas.
c.
Plásmidos de Resistencia (factores R) – Estos plásmidos acarrean
genes de resistencia a los antibióticos.
i) Origen
El origen de los factores R no se conoce. Es probable que hayan
evolucionado con otros propósitos y que el advenimiento de la era de los
antibióticos haya proporcionado una ventaja selectiva para su amplia
diseminación.
|
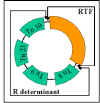 Figura 11 Estructura del Plásmido R
Figura 11 Estructura del Plásmido R |
ii)
Estructura
Los plásmidos R son plásmidos conjugativos en los
cuales los genes para la replicación y la transferencia se localizan en
una parte del factor R y los genes de resistencia están localizados en
otra parte del mismo, como se ilustra en la Figura 11.
RTF (Resistance Transfer Factor) – acarrea los genes de
transferencia.
Determinante R – acarrea los genes de resistencia. Los genes de
resistencia frecuentemente forman parte de transposones.
Mecanismo de acción
de los genes de resistencia
a) Modificación (detoxificación) de antibióticos - ej. la enzima β-lactamasa
b) Alteración del sitio blanco -
ej. resistencia a la
estreptomicina
c) Alteración de la incorporación- resistencia a la Tetraciclina
d) Substitución de una ruta sensible -
ej. Una nueva ruta de
síntesis del ácido fólico como mecanismo de resistencia a las drogas
conocidas como sulfas.
|
|
|
 Regreso
a la Sección de Bacteriología de Microbiología e Immunología On-line Regreso
a la Sección de Bacteriología de Microbiología e Immunología On-line
El mantenimiento de ésta página es conducido por
Richard Hunt
|
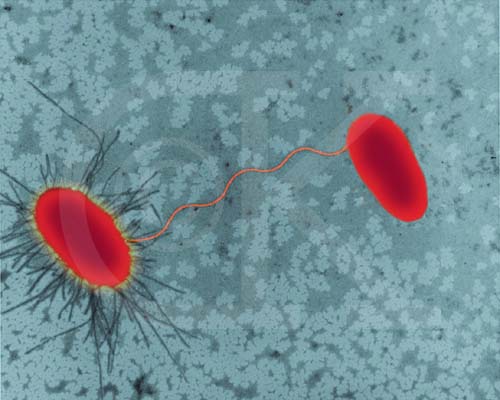 E. coli
(bacilo procariote) cepas llevandoa cabo la conjugación.
Una cepa tiene fimbrias
©
Dr
Dennis Kunkel,
University of Hawaii. Used with permission
E. coli
(bacilo procariote) cepas llevandoa cabo la conjugación.
Una cepa tiene fimbrias
©
Dr
Dennis Kunkel,
University of Hawaii. Used with permission
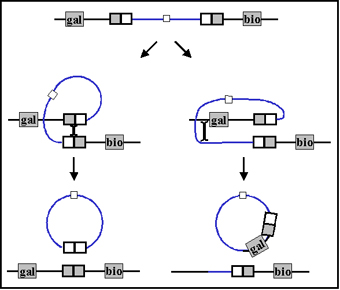 Figura 4. El mecanismo de la transducción especializada.
Figura 4. El mecanismo de la transducción especializada. 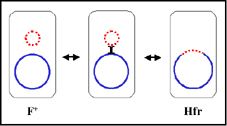 a
a
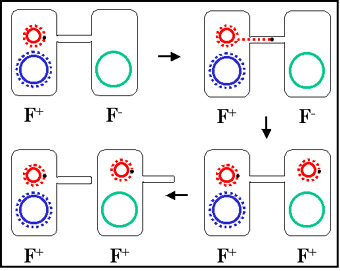 Figura 6 Mecanismo de los cruces F+ x F-
Figura 6 Mecanismo de los cruces F+ x F-  Figura 8 El mecanismo de cruces F" x F-
Figura 8 El mecanismo de cruces F" x F- 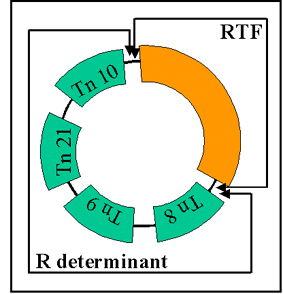 Figura 11 Estructura del Plásmido R
Figura 11 Estructura del Plásmido R 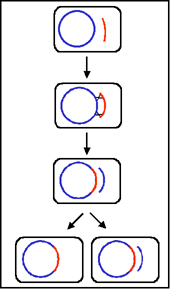

{kind=link}
{kind=link}
{kind=link}