|
x |
x |
|
 |
 |
|
INFECTIOUS
DISEASE |
BAKTERIYOLOJİ |
İMMÜNOLOJİ |
MYCOLOGY |
PARASITOLOGY |
VIROLOGY |
|
|
İMMÜNOLOJİ- BÖLÜM DÖRT
İMMÜNGLOBULİNLERİN GENETİĞİ
Gene Mayer, Ph.D
Emertius Professor of Pathology, Microbiology and Immunology
University of South Carolina
Çeviri:
Doç. Dr. Erkan Yula
İzmir, Katip Çelebi Üniversitesi, Tıp Fakültesi, Tıbbi Mikrobiyoloji
Anabilim Dalı
|
|
ENGLISH |
|
FRANCAIS |
|
PORTUGUES |
|
SHQIP |
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|

 |
|
Logo image
© Jeffrey Nelson, Rush University, Chicago, Illinois and
The MicrobeLibrary |
|
|
|
|
|
ÖĞRENİM HEDEFLERİ
İmmünoglobülin gen ailesinin organizasyonunu ve ekspresyonunu tanımlamak.
Antikor çeşitliliğinin nedenini açıklamak
|
TARİHÇE
Aminoasit sekans verileri, tek bir C bölgesi’nin birçok farklı V
bölgeleriyle bağlantılı olabileceğini göstermiştir. Ayrıca tek bir idiotipin
farklı C bölgeleriyle (IgM ve IgG) bağlantılı olabileceği de gösterilmiştir. Bu
bilgiler ışığında immünglobulin molekülünün bu iki bölgesi birbirinden farklı
genlerce kodlandığı düşünülmüş; V ve C bölgelerini kodlayan genler immünglobulin
yapılmadan önce bir şekilde yapıya katılmış olabildiği kanatine varılmıştı (örn
tek bir polipeptid için iki gen bölgesi bulunması gibi). Bu durum devrim
niteliğindeydi ve rekombinant DNA teknolojisindeki gelişmeler sonucunda doğru
olduğu kanıtlanmıştır.
İmmünglobulinin hafif ve ağır zincirleri her biri farklı
kromozomlarda bulunan üç farklı gen ailesince kodlanır; ağır zincir için bir
tane, hafif zincir tiiplerinin her biri için bir tane. Bu gen ailelerinin
herbiri, birkaç V bölgesi geni ve bir veya birden fazla C bölgesi geni içerir. V
ve C bölgesi genleri birbirine komşu değildir.
|
|
ANAHTAR KELİMELER
V geni
C geni
J bölgesi
D bölgesi
Leader
Enhancer
Promotor
Antikor çeşitliliği
Germ line theorisi
Somatik mutasyon theorisi
N bölge insersiyonları
Birleşme çeşitliliği
Kombinasyonal ilişki
Multispecifisite
Klonal seleksiyon
|
HAFİF ZİNCİR GEN AİLESİ
Gen organizasyonu
Şekil 1’de diferansiye olmamış hücrelerin kappa ve lambda hafif zincir
genlerinin düzenlenimi gösterilmektedir.
-
Lambda Hafif Zincir
Lambda gen ailesi 4C bölgesi genleri tarafından oluşturulur, lambda
zincirinin her bir subtipi için 1 adet ve yaklaşık olarak 30 V bölgesi
geni. Her bir V bölgesi genleri iki ekson tarafından oluşturulur; biri
(L) lider bölgeyi, diğeri (V) değişken bölgenin çoğunu kodlar. Her bir C
bölgesinin üzerinde ek olarak J (joining) eksonu bulunmaktadır. L, V, J
ve C eksonları intronlarca bölünmüştür (kodlanmayan sekansların arasında).
|
 Şekil 1
Şekil 1
Kappa ve lamda hafif zincir genlerinin germ line ve farklılaşmamış hücre
düzeyinde organizasyonu
|
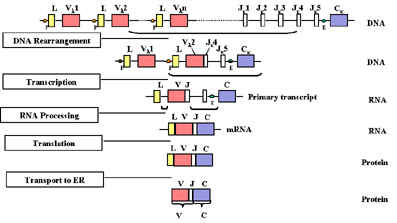
Gen ekspresyonu ve yeniden
düzenlenmesiBir hücre hafif zincir üretecek olan matür
B lenfosite diferansiye olduğunda, çeşitli genlerin (eksonlar)
yeniden düzenlenimi gerçekleşir ve genler Şekil 2’de gösterildiği
gibi eksprese olmaya başlar.
ir hücre, hafif zincir üretecek olan bir B lenfosite dönüşmeye
başladığı zaman, V geninin J bölgelerinden birinin yanına
getirilmesi gibi DNA düzeyinde genlerin yeniden düzenlenimi söz
konusudur. Bu durum V ve J bölgeleri arasındaki intronun
çıkarılmasını içeren bir rekombinasyon sayesinde olur. Hangi V
geninin seçileceği tam olarak tesadüfi değildir; J bölgesine en
yakın V geninin kullanımı önceliklidir. Fakat zamanla tüm V genleri
kullanılabilir; böylece V geni ve J bölgesinin tüm kombinasyonları
oluşturulabilir. DNA yeniden düzenleniminin bir sonucu olarak gen
transkripsiyona hazır hale gelir çünkü V geniyle bağlantılı olan
promoter(P), J ve C bölgeleri arasındaki intronda bulunan enhancer
(E) ile yakın hale getirilir. Transkripsiyon promoter bölgesinden
başlar; L-V, J-C arası intronların sekanslarını içerdiği gibi L,V,J,
C bölgesi sekanslarını da içeren bir pre-mRNA yapılır (Şekil 2)
Bu pre-mRNA nükleusta işlenir ve artakalan intronlar atılır.
Sonuçta oluşan mRNA L, V, J ve C eksonları yan yanadır. mRNA
sitoplazmada translasyona uğrar ve protein endoplazmik retikulum
lümenine transporte edildiğinde lider zincir kaldırılır. Endoplazmik
retikulumda hafif zincir, ağır zincir ile birleştirilir ve
immünglobulin normal sekretuar proteinler yoluyla sekrete edilir.
Matür hafif zincirin V bölgesi, V geni sekansları tarafından
kodlanır ve J bölgesi sekansları ve C bölgesi, C geni sekansındaki
tarafından kodlanır.
|
 Şekil 2
Şekil 2
Hücrenin olgun B hücreye farklılaşmasında hafif zincir yapılacaktır ve
çeşitli genlerde (ekzon) yeniden düzenlenim bulunmaktadır.
|
AĞIR ZİNCİR GEN AİLESİ
Germ line gen
organizasyonu
Ağır zincir genlerinin düzeni Şekil 3‘te gösterilmiştir.
Ağır zincir gen ailesinde, biri immünglobulinlerin herbir sınıf ve altsınıfı
için olmak üzere, birden fazla C geni bulunur. Her bir C geni birçok ekson
tarafından oluşturulur, herbir domain için bir; menteşe bölgesi için bir
diğer ekson bulunmaktadır. Ağır zincir gen ailesinde birçok V geni bulunur;
her biri lider zincir ve V eksonundan oluşmuştur. J eksonlarına ek olarak
ağır zincir gen ailesi ayrıca D (diversity) eksonları olarak adlandırılan
birçok ek ekzonu da içerir. Şekil 3’te gösterildiği gibi tüm eksonlar
intronlarca parçalara ayrılmıştır.
|
 Şekil 3
Şekil 3
Birkaç J ekzonuna ek olarak, ağır zincir gen ailesi aynı zamanda D
(diversity) olarak adlandırılan birkaç ek ekzon içermektedir. Tüm
ekzonlar intronlar ile ayrılmıştır.
|
Gen yeniden
düzenlenmesi ve ekspresyonu
Bir hücre ağır zincir üretecek
olan matür B hücreye farklaştığı zaman, eksonlar arasında çeşitli
genlerin yeniden düzenlenimi olur ve Şekil 4 ve 5’te gösterildiği gibi
bu gen eksprese olmaya başlar. Bir hücre ağır zincir üretebilen bir B
lenfosite dönüşeceğinde, DNA basamağında iki adet yeniden düzenlenime
uğrar. İlki, D bölgelerinden biri J bölgelerinden birinin yanına
getirilir ve sonrasında V genlerinden biri yeniden düzenlenime uğramış
DJ bölgesinin yanına getirilir. Bu durum V, D ve J bölgeleri arasındaki
intronların çıkarılmasını içeren iki rekombinasyon basamağı ile
gerçekleşmektedir. Hafif zincirlerde olduğu gibi ağır zincir V geni
seçilimi de tam olarak randomize değildir fakat sonuçta tüm V genleri
kullanılabilir. Bu DNA rearanjmanlarının bir sonucu olarak gen,
transkripsiyonel olarak aktif hale gelir çünkü V geniyle bağlantılı olan
bir promoter (P), J ve Cmu bölgeleri arasındaki intronda lokalize olan
enhancer (E) bölgesine yakın konuma getirilir. Promoter bölgesinde
transkripsiyon başladığında L-V, J-Cmu, Cmu ve Cdelta arasındaki intron
sekanslarını içerdiği gibi; L, V, D, J, Cmu ve Cdelta bölgelerinin
sekanslarını içeren bir pre-mRNA yapılır (Şekil 4). Pre-mRNA çekirdekte
işlenir ve C genindeki eksonlar arasında artakalan intronlar
uzaklaştırılır (bakınız şekil 5).
|
| |
Pre-mRNA iki şekilde işlenebilir; birincisi VDJ
bölgesini Cmu geninin
yanına getirmek; bir diğeri VDJ bölgesini Cdelta geninin yanına
getirmektir. Sonuçta mRNA, L, V, D, J ve Cmu veya Cdelta
eksonlarını ardısıra içerir ve sırasıyla mu ve
delta zinciri kodlanır. mRNA sitoplazmada translasyona uğrar
ve proteinler endoplazmik retikulum lümenine transporte edildiğinde
lider kısım uzaklaştırılır. Ağır zincir endoplazmik retikulumda
hafif zincirle birleştirilir ve immünglobulin sekretuar proteinler
yoluyla sekrete edilir. Matüre olmuş ağır zincirin V bölgesi, V
geni, D bölgesi, J bölgesi tarafından kodlanır ve C bölgesi C
genindeki sekanslarca kodlanır.
|
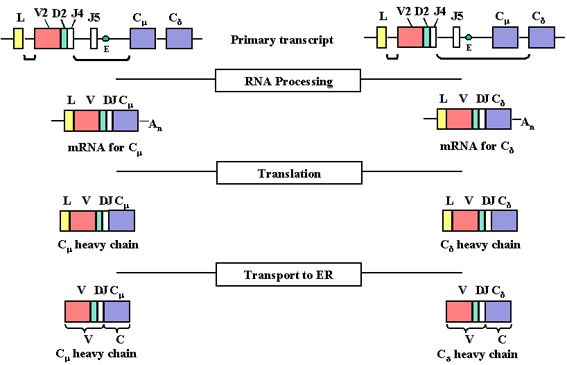
 Şekil 4
Şekil 4
Promoterden transkripsiyon başladığında pre-mRNA yapılır. Bu V, D, J Cμ
ve Cδ bölgerei yanı sıra L ve V arasında, J ve Cμ arasında ve Cμ ve Cδ
arasında intron sekansları içermektedir.
 Şekil 5
Şekil 5
pre-mRNA nükleusta işlenir (uç uca eklenen) ve C genindeki ekzonlar
arasndaki kalan kısım uzaklaştırılır.
|
DNA yeniden düzenlenme MEKANİZMASI
Birbirine komşu
V, J ve D eksonlarında, rekombinasyon fonksiyonları olan rekombinasyon
sinyal sekansları (RSS) olarak adlandırılan özgün sekanslar bulunmaktadır.
Her bir rekombinasyon sinyal sekansı korunmuş bir nanomer ve heptamer
içerir; bunlar 12-23 baz çifti ile birbirinden ayrılmıştır (Şekil 6). Bu 12
ve 23 baz çifti içeren bölgeler DNA heliksinin bir veya iki dönüşünden
sorumludur. Rekombinasyon sadece bir ve iki dönüşü arasında olur. λ hafif
zincirlerde J ekzondan yukarı doğru 1 dönüş sinyal vardır, Vlambda.
‘dan aşağı doğru 2 dönüş sinyal bulunmaktadır. κ hafif zincirlerde Vkappa genden
aşağı doğru 1 dönüş sinyal vardır, J ekzondan yukarı doğru 2 dönüş sinyal
bulunmaktadır. Ağır zincirlerde, D ekzonun her bir tarafı için
1 dönüş
sinyal vardır ve V genindan aşağı doğru 2 dönüş sinyali ve J ekzondan yukarı
doğru 2 dönüş sinyali bulunmaktadır. Böylelikle doğru rekombinasyonun
sağlanması garantilenmiş olmaktadır. Rekombinasyon hafif zincir için V ve J
bölgeleri arasındaki intronların veya ağır zincir için V, D, J bölgeleri
arasındaki intronların çıkarılmasıyla sonuçlanır. Rekombinasyon olayı Rag-1
ve Rag-2 adlı iki protein tarafından katalize edilir. Bu iki proteini
kodlayan genlerdeki mutasyon ağır kombine immün yetmezlikle sonuçlanır (T ve
B lenfositlerin ikisinin birden eksikliği), çünkü bu proteinler ve
rekombinasyon sinyal sekansları (RSS) B ve T lenfosit reseptörlerinin
oluşumuna katılmaktadırlar.
|
 Şekil 6
Şekil 6
Komşu V, J ve D ekzonları benzersiz diziye sahiptir ve rekombinasyon
sinyal sekansları (RSS) olarak adlandırılır. Her bir RSS korunmuş
nonamer ve korunmuş heptamerden oluşmaktadır ve her bir 12 veya 23 baz
çifti ile ayrılmışlardır. |
İMMÜNGLOBULİN GEN AİLESİNDE GEN
EKSPRESYON DÜZENİ
Özgün bir B lenfosit sadece tek tip hafif zincir ve tek sınıf ağır zincir
üretir. (Tek istisnası matür B lenfosit μ ve δ ağır zincirin her ikisini de
üretebilir fakat μ ve δ zincirlerinde aynı VDJ bölgeleri bulunduğu sürece
antikor spesifitesi de aynıdır). Herhangi bir B lenfosit hem anneden hem de
babadan gelen immünglobulin genlerini kodlayan kromozomları içerir; bir
hücre, belli bir tip hafif zincir ve belli bir sınıf ağır zincir
üretileceğinden emin olmak için immünglobulin gen ekspresyonunda belli bir
yol izlemelidir. Şekil 7 ve 8’de immünglobulin genlerinin hangi sıra ile
eksprese edildiği gösterilmektedir.
|
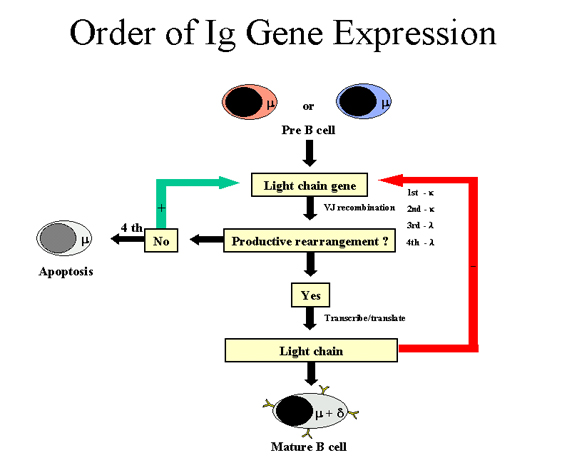
 Şekil 7
Şekil 7
Ig gen ekspresyon sırası- Ağır zincir
 Şekil 8
Şekil 8
Ig gen ekspresyon sırası- Hafif zincir
|
Ağır zincir (Şekil 7)
Bir hücre ilk olarak ağır zincir genlerini yeniden düzenlemeye
çalıştığında; bazı hücrelerde maternal kromozom, bazılarında paternal
kromozom seçilir. Eğer bu yeniden düzenlenim başarılı olup ağır zincir
yapılırsa, artık ağır zincir genlerinde bir daha düzenlenim yapılmaz. Fakat
diğer taraftan, eğer ağır zincir genlerindeki bu ilk yeniden düzenlenim
başarılı olmazsa (örn hiç ağır zincir yapılamazsa), hücre diğer kromozom
üzerinden bunu yapmaya çalışır. Eğer hücre ikinci kez de ağır zincir
genlerini yeniden düzenlemeyi başaramazsa, bu hücre elimine edilecektir.
Kappa hafif zincir (Şekil
8)
Bir hücre ağır zincir genindeki yeniden düzenlenim başarılıyla
tamamladığında, ardından kappa hifif zincir genlerinden birinin yeniden
düzenlenimi başlar. Maternal veya paternal kappa zincir genlerinden
hangisinin seçileceği randomize bir olaydır. Eğer yeniden düzenlenim
başarılı olmazsa (örneğin fonksiyonel bir kappa zincir üretemiyorsa), o
zaman diğer kromozomdaki kappa genlerini düzenlemeye çalışır. Eğer bir hücre
kappa hafif zincir genini başarılı bir şekilde yeniden düzenlenime
uğratabilirse, kappa hafif zinciri olan bir immünglobulin üretebilen bir B
lenfosit olmuş olur.
|
| |
Lambda hafif zincir
(Şekil 8)
Eğer bir hücre her iki kappa hafif zincir genini yeniden
düzenlemede başarısız olursa, lambda hafif zincir yapmayı dener.
Maternal veya paternal lambda hafif zincirlerinden hangisinin
seçileceği randomize bir olaydır. Eğer bu yeniden düzenlenim
başarısız olursa (örn fonksiyonel bir lambda hafif zincir üretemezse),
diğer kromozomdaki lambda genini yeniden düzenlenime sokar. Eğer bir
hücre lambda hafif zincir genini başarılı bir şekilde yeniden
düzenlenime uğratırsa, lambda hafif zinciri olan immünglobulin
üretebilen bir B lenfosit olmuş olur.
|
| |
İmmüngloobulin gen ailesi sistemli bir sıra ile yeniden
düzenlenimi şu sorulara açıklık getirir:
-
Neden tek bir B lenfosit sadece belli bir tip
hafif ve ağır zincir içeren immünglobulin üretebilir?
-
Niçin belli bir B lenfosit, beli bir spesifitede
antikor üretebilir?
-
Niçin bir immünglobulin molekülü seviyesindeki
immünglobulin allotipi alelik çıkarıma uğrar fakat allotiplerin
ko-dominant ekspresyonu toplu olmaktadır?
|
 Şekil 9
Şekil 9
Şuanki konsepte göre antikor çeşitliliğinin kökeni
|
ANTİKOR ÇEŞİTLİLİĞİNİN KÖKENİ
Antikor çeşitliliği, bir organizmanın üretebilmesi mümkün olan spesifik
antikorların toplamını ifade eder. 107 - 108 farklı antikor molekülü
üretebildiğimiz tahmin edilmektedir. İmmünolojinin önemli sorularından
biri de bu kadar farklı antikor molekülünü nasıl yaptığımızdır. Antikor
çeşitliliğini açıklamaya çalışan teoriler iki majör kategoriye ayrılır.
Germ Line Teorisi
Bu teori, üretilecek her bir antikor molekülü için farklı bir V
gen bölgesine sahip olduğumuzu ifade eder.
Somatik Mutasyon Teorisi
Bu teori ise sadece bir veya birkaç V bölgesigenine sahip
olduğumuzu ve çeşitliliğin bu genlerdeki somatik mutasyonlardan
kaynaklandığını söyler.
Şu Anki Durum
Bizim şu anki düşüncemiz ise hem germ line teorisinin hem de
somatik mutasyon teorisinin de geçerli olduğudur.
Antikor çeşitliliğinin şu mekanizmalara bağlı olduğu düşünülüyor:
1. V gen sayısının çok fazla olması
a ) 30 lambda V geni
b) 300 kappa V geni
c) 1000 ağır zincir V geni bulunmaktadır.
2. V-J ve V-D-J birleşmesi
Hafif zincir V geni ve J bölgesi veya ağır zincir V geni, D
ve J bölgelerinin biraraya geldiği bölge üçüncü hipervariable
bölgedir. Hangi V ve hangi J veya D bölgelerinin bir araya
geleceği randomize olarak seçildiğinden, V-J ve V-D-J
birleşmesinden çok fazla çeşitlilik üretilebilmektedir.
|
| |
3. Birleşme çeşitliliği (V-J ve V-D ve D-J rekombinasyon hataları)- (Şekil
9) V-J ve V-D-J arasındaki rekombinasyon her zaman mükemmel bir
biçimde gerçekleşmemektedir ve bu çeşitlilik rekombinasyondaki hatalar
sayesinde de artar; örneğin V bölgesini J veya D bölgesinin yanına
getirilmesi veya D bölgesini J bölgesinin yanına getirilmesi. Bu
hataların V-J ve V-D-J birleşmesiyle çeşitliliği üç kat arttırdığı
tahmin edilmektedir. Bu mekanizmayla oluşturulan çeşitlilik üçüncü
hipervariable bölgede gerçekleşmektedir ve böylelikle direkt olarak
antikorun birleşme noktasını etkilenmektedir. 4. N bölgesi insersiyonu
D ve J segmentleri arası birleşme noktasında terminal transferaz
enziminin katalizlediği bir nükleotitd insertion serisi sıklıkla vardır.
Terminal transferaz, DNA’da kalıba ihtiyaç duymadan nükleotid
polimerizasyonunu randomize olarak katalizler. Bu durum üçüncü
hipervariable bölgede daha fazla çeşitliliğe neden olur. 5.Somatik
mutasyon V geninde somatik mutasyonlar olduğuna dair kanıtlar
bulunmaktadır; özellikle ikinci hipervariable bölgeyi kodlayan bölgede.
Böylelikle, somatik mutasyonlar muhtemelen belli bir ölçüye kadar
antikor çeşitliliğine katkıda bulunuyor olabilir. 6. Kombinasyonel
ilişki
Herhangi bir özgün B lenfosit mümkün olan herhengi bir hafif ve ağır
zinciri yapabilme potansiyeline sahiptir. Böylelikle hafif ve ağır
zincirin farklı kombinasyonları B lenfosite daha fazla çeşitlilik
kazandırır. 7.Multispesifite
Benzer yapıdaki antikorların antijenik determinantları arasındaki çapraz
reaksiyondan dolayı bir antikor sıklıkla birden fazla antijenle
reaksiyona girebilir. Bu durum multispesififte olarak adlandırılır.
Multispesifite ayrıca antikor çeşitliliğine katkı sağlar.
Bu mekanizmanın bu kadar fazla çeşitliliğe nasıl katkı sağlayabildiği
aşağıda gösterilmektedir:
|
| |
|
|
B hücre Reseptörü (immünoglobülin) |
|
Ağır |
Kappa |
|
V gen segmenti |
1000 |
300 |
|
D gen segmenti |
15 |
- |
|
J gen segmenti |
4 |
4 |
|
N bölge insersiyonu |
++ |
- |
|
Birleşme çeşitliliği |
+++ |
+ |
|
Somatik mutasyon |
+ |
+ |
|
Kombinasyonel ilişki |
V x D x J
1000 X 15 X 4 |
V x J
300 x 4 |
|
Toplam |
6 x 104 |
1.2 x 103 |
|
|
|
|
Kombinasyonel ilişki |
7.2 x 107 |
|
| |
Bu hesaplamalarda lambda hafif zincir, somatik mutasyon
birleşme çeşitliliği, N bölgesi insertionları veya multispesifitenin katkısı
göz ardı edilmiştir. Hafif ve ağır zincirlerde gen yeniden düzenlenim işlemi
ve bu zincirlerin kombinasyonel ilişkisi, B lenfosit gelişimi boyunca sürer
ve antijenden bağımsızdır. Tüm muhtemel özgül antikor ekprese eden B
lenfosit klonları gelişim boyunca üretilmeye devam eder ve antijen basitçe
uygun reseptöre sahip olan klonları seçmektedir. Seçilen klonlar aktive olur,
proliferasyona uğrar ve antikor sekrete eden plazma hücrelerine
farklılaşırlar.
ANTİJEN İÇİN T
LENFOSİT RESEPTÖRÜ
T lenfositler yüzeylerinde ayrıca antijen için reseptör
taşırlar. Bu reseptör bir immünglobulin molekülü değildir fakat
immünglobulinlerle analog yapıda sabit ve değişken bölgeleri olan, iki
farklı polipeptid zinciri tarafından oluşturulur. T lenfosit reseptör
çeşitliliği, antikor çeşitliliğinde tarif edildiği gibi aynı şekilde
gerçekleşir (örneğin gen segmentlerinde VJ ve VDJ bağlantısı ve
kombinasyonel ilişki). Fakat T lenfositlerde hiçbir somatik mutasyon
gözlenmemiştir.
|
|
|
 Mikrobiyoloji ve İmmünoloji On-line, İMMÜNOLOJİ Bölümüne Dönünüz
Mikrobiyoloji ve İmmünoloji On-line, İMMÜNOLOJİ Bölümüne Dönünüz
This page last changed on
Sunday, March 27, 2016
Page maintained by
Richard Hunt
|
 Şekil 2
Şekil 2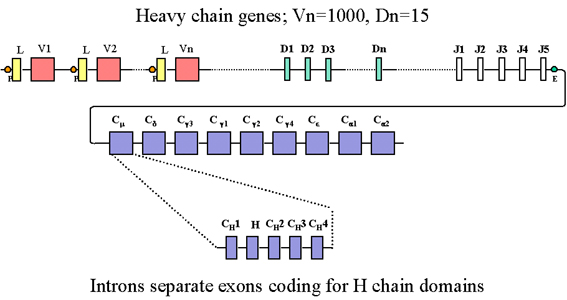 Şekil 3
Şekil 3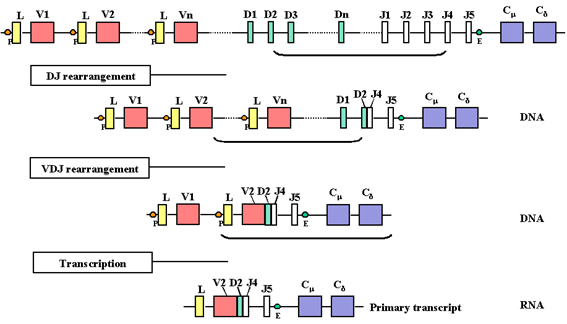 Şekil 4
Şekil 4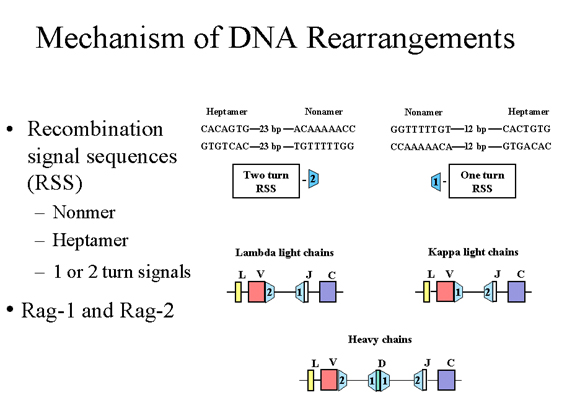 Şekil 6
Şekil 6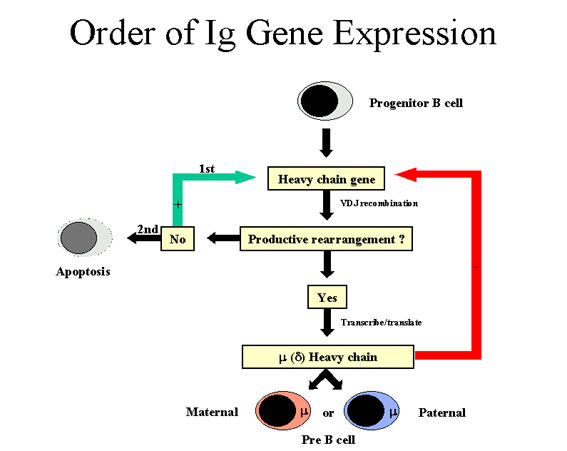 Şekil 7
Şekil 7 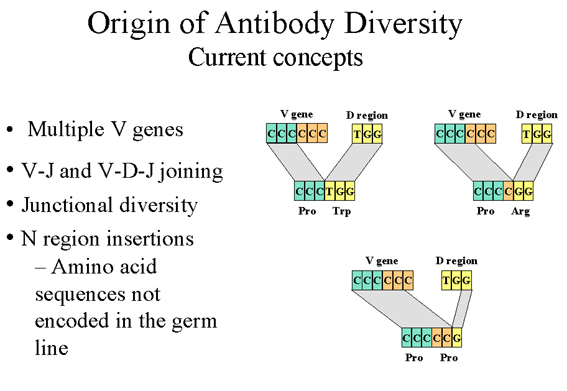 Şekil 9
Şekil 9