|
xx |
x |
|
 |
 |
|
INFECTIOUS
DISEASE |
BAKTERIYOLOJİ |
İMMÜNOLOJİ |
MYCOLOGY |
PARASITOLOGY |
VİROLOJİ |
|
|
VİROLOJİ- BÖLÜM BEŞ
VİRAL GENETİK
Dr Margaret Hunt
Professor Emerita
University of South Carolina School of Medicine
Çeviren.
Prof. Dr. Selçuk Kaya
İzmir Katip Çelebi Üniversitesi, Tıp Fakültesi
|
|
SPANISH |
|
ENGLISH |
|
ALBANIAN |
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|

 |
|
|
|
|
|
ÖĞRETİM HEDEFLERİ
Hayvan viruslarının genetiğine giriş
|
GENEL
Virüsler hızlı bir şekilde büyürler ve hücre başına genelde
büyük miktarda virion soyları bulunur. Bu yüzden kısa bir zaman dilimi içinde
mutasyon olma şansı daha fazladır.
Viral genomun doğası (RNA veya DNA; bölünmüş yada bölünmemiş) virüslerin
genetiğinde önemli bir rol oynar.
Virüsler mutasyon veya yeniden birleşme (rekombinasyon) nedeniyle genetik olarak
değişebilirler
MUTANTLAR
Köken
Spontan mutasyonlar
Bunlar viral üreme sırasında doğal olarak ortaya çıkar,
örnegin genom-üreme polimerazı esnasındaki hatalar sonucu veya bazların
tautomeric (iki izomer arasındaki dinamik denge) formlarının birleşmesi
sonucu.
DNA virüsleri RNA virüslerine göre genetik olarak daha stabil olmaya
meyillidir. Konak hücrede DNA onarımı için hata düzeltme mekanizmaları
bulunurken, RNA için muhtemelen bu yoktur. Bazı RNA virüsleri kayda
değer bir bicimde doğada sabit değerdedir.
Muhtemelen bu virüslerde diğer RNA virüsleri ile aynı oranda yüksek
mutasyon oranı bulunur ancak bulaşma ve üremeye yüksek adaptasyonları
sayesinde küçük değişimler ana virüs (yabani-tür, brüt) ile mücadelede
başarısız olur.
Mutasyonlar fiziksel veya
kimyasal vesileler ile oluşur
Kimyasal
Bazlara direk etki eden etmenler mesela nitröz asit
Endirek etmenler örnegin baz analogların normal bazlara göre daha
sık yanlış eşleşmesi sonucu oluşan mutasyonlar
Fiziksel
X - ışını veya ultra viole ışınlar gibi etmenler
Mutasyon tipleri
Mutantlar nokta mutant olabilir (bir bazın başka biri
tarafından yer değiştirmesi) veya ekleme/silme mutantlar.
Mutant Virüslerde
görülen Fenotipik degisim cesitlerinin örnekleri
(fenotip= bir organizmanın gözle görünen özellikleri)
Koşullu ölümcül mutantlar
Bu mutantlar diğerlerinden (yabani tipdeki virüsler her
iki şart altinda da coğalırlar) farkli olarak sadece bazı sartlar
altinda çoğalırlar.
Örnek olarak ısıya duyarli olan mutantlar (ts) -
Bunlar 31 C derece gibi düsük isilarda büyürler ancak örnegin 39 C
derecede buyumezler, yabani tipler 31 ve 39 C dereceler arasında
büyürler. Öyle görünüyorki bunun sebebi genellikle değiştirilmiş
proteinin yüksek sıcaklıklarda fonsiyonel bir adaptasyon
sağlayamamasıdır.
Örnek olarak konak çeşiti - Bu mutantlar sadece bazı
hücre tiplerinde büyürler ancak yabani çeşitteki virüsler büyürler -
bu çeşit mutantlar viral enfesksiyonda konak hücrenin rölünü
incelemek icin bazı yollar sağlarlar.
Plak boyutu
Plaklar yabani virüs çeşitlerine göre daha büyük veya
daha küçük olabilirler, bazen bu tip mutantlar değişmiş patojenlik
gösterirler.
İlaç direnci
Anti-viral etmenlerin geliştirilmesinde bu önemlidir -
Mutantlarda ilaca karşı direncin artma ihtimali sürekli göz önünde
bulundurulmak zorundadır.
Enzim eksiği mutantlar
Bazi viral enzimler her zaman gerekli değildir ve bu
yüzden enzim eksiği mutantların izole edilmesi uygulanabilir; örneğin
herpes simpleks virüsü timidin kinaz her zaman doku kültüründe gerekli
değildir ancak sinirsel hücrelerin enfeksiyon olmasında önemlidir.
"Sicak" Mutantlar
Bunlar yabani virüs tiplerine göre yüksek sıcaklıklarda
daha iyi büyürler. Bunlar daha ölümcül olabilirler çünkü konağın yüksek
ateşi bu mutantlar üzerinde çok küçük etkisi olur ancak yabani tipdeki
viryonların üremesini yavaşlatabilir.
Zayıflatılmış mutantlar
Bircok viral mutantlar ana virüslere göre daha hafif
semptomlara (veya sıfır semptoma) sebep olur - bunlara zayıflatılmış
denir. Bunlar aşı gelistirilmesinde potansiyel bir role sahiptir ve aynı
zamanda ana virüslerin neden zararlı olduğunu belirlemede yararlı bir
araçtırlar.
|
 Şekil 1
Şekil 1
Kopya secim rekombinsayonu |
GENETİK MATERYAL DEGİŞİMİ
Rekombinasyon
Iki genom arasında genetik bilgi degisimi.
"Klasik" rekombinasyon
Bu nukleik asitlerdeki kovalent baglarin kirilmasi,
genetik bilgilerin degisimi ve kovalent baglarin yeniden olusmasini
kapsar.
Bu cesit kirilma/ekleme rekombinasyonu DNA dirüsleri
veya DNA fazı (retrovirusler)na sahip RNA larda yaygindir. Konak hücre
DNA icin rekombinasyon sistemlerine sahiptir.
RNA virüslerinde bu cesit rekombinasyonlar cok seyrektir
( Muhtemelen RNA rekombinasyonu icin konak enzimleri bulunmamaktadir).
Picorna virusler cok kücük oranda rekombinasyon verimi gösterirler.
Mekanizma standart DNA mekanizmasından farklıdır ve muhtemelen içinde
polimerazın RNAyı kopyalarken taslakları yer değiştirdiği bir "kopya
secim" mekanizması türüdür (Şekil 1) .
Rekombinasyon korona viruslerde de yaygındır - Burada da
mekanizma DNA’nın durumundan farklıdır ve muhtemelen virus içinde
RNA’nın olağan dışı sentezinin sonucudur.
Simdiye kadar negatif sarmallı RNA virüslerindeki
rekombinasyonun gecerli vrüslerin artışına neden olduguna dair bir kanit
bulunamamistir (Bu virüslerde, genomik RNA nucleokapsidlerde paketlenir
ve gen eşleşmesi için hazır durumda değildir)
|
 Şekil
2 Şekil
2
Markır kurtarma |
Rekombinasyon tekniklerinin
kullanım cesitleri
-
Genomların haritalandırılması ( iki genin
birbirinden daha uzak olmasi ikisi arasindaki rekombinayonun olma
olasılığını artırır.)
-
Markır kurtarma - Yabani virüs cesitlerine ait DNA
parcaları (fragmentleri) yabani tipde virüs olusturmak icin mutant
virüslerle rekombine edilebilinir. Bu, genomdaki belirli bir bölgeye
gen fonksiyonu verilmesi icin bir yol saglar.
Hatta bu bir gene yabanci bir materyalin yerlestirlmesi
icin de bir yol saglar (Şekil 2).
Rekombinasyon virüslere aynı tip virüslerden ve nadiren aynı tip olmayan
virüslerden veya konak genomdan genetik bilginin alınmasına olanak
sağlar (bazi retro virüslerde oldugu gibi - retrovirüslere bakınız)
|
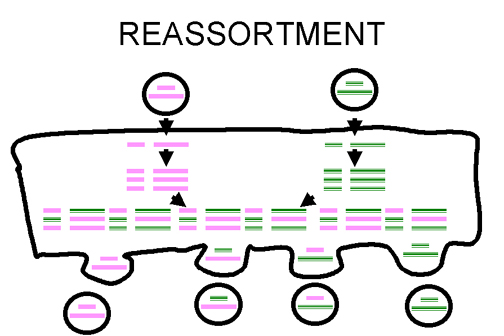
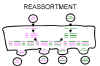 Şekil 3
Şekil 3
Segmentli viruslarda virüs genomunun yeniden dizilimi |
Yeniden Dizilim
Eger bir virüsün segmentli genomu var ise ve bu virüsun
iki degiskeni bir hücreye bulaşırsa, bazı segmentleri bir ebeveynden ve
bazi segmentleri de diger ebeveynden olmak üzere viryon soylarinin
oluşmasını sağlar.
Bu etkili bir prosesdir - fakat segmentli virüslere
uygalanabilir - şimdiye kadar segmenti genomlarla karakterize edilmis
insan virüsları sadece RNA virüsleridir ve bunlara örnek olarak
orthomyxovirüsler, reovirüsler, arenavirüsler, bunya virüsleri
verilebilinir.
Yeniden dizilim dogada özgün reassortantların (iki ya da daha fazla
benzer virüsten elde edilen genetik materyal) olusumunda önemli bir rol
oynayabilir ve labaratuar deneylerinde de fayda sağlar (Şekil 3).
Genomlarin farklı segmentlerine fonksiyon yüklenmesinde
de kullanılmıştır. Örnegin, yeniden dizilmis bir virüsde bir segment A
virüsünden geliyorsa ve gerisi B virüsünden geliyorsa, hangi
özelliklerin A virüsüne ve hangilerinin B virüsüne benzedigini
görebiliriz.
Yeniden dizilim klasik olmayan bir rekombinasyonun türüdür
|
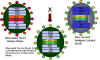 Şekil 4
Şekil 4
Zayıflatılmış grip virüsü soyu ve zayıflatılmış grip aşısı
formasyonundaki yeni kuvvetli bir soy arasında genlerin yeniden dizilimi
Adapted from: Treanor JJ Infect. Med. 15:714 |
Uygulamalı genetik
Yukarda tartışılan prensiplerin bir kismını içeren grip virüsü icin
kullanılan Flumist (LAIV, Haziran 2003 onaylanmistir) denilen bir aşı
vardir. Aşı üclüdür - 3 soy grip virüsü icerir.
Virüsler 25 ve daha soğuk C derecede üst solunum yollarında da
buyuyen soğuğa adapte olmuş soylardır. Virüsler sıcaklığa duyarlıdır
ve sıcak alt solunum yollarında yavaş büyür. Virüsler zayıflatılmış
suşlardır ve yabani tipdeki virüslerden daha az patojeniktir. Bunun
nedeni cesitli genom segmentlerinde olan birden fazla degisikliktir.
Grip virüsü yüzey proteinlerine (HA - hemagglutinin ve NA -
neuraminidase) karşı antikorlar enfeksiyondan korunma için önemlidir.
HA ve NA yıldan yıla değişir. Aşı teknolojisi zayıflatılmış, soğuğa
adapte olmus virüs ve yeni dönemde problem olacak virüslerin HA ve
NA kodlu segmentlerini yeniden dizilişi teknikleri kullanarak 6 gen
segmentli reassortant virüsleri elde eder.
Aşı yasayan bir aşıdır ve burun içi yoluyla sprey olarak verilir
ve mukozal ve sistemik bağışıklığı başlatır.
Yakin gecmiste yaşayan zayıflatılmış bir reassortant rotavirüs (RotaTeq
from Merke) aşısı olarak onaylanmıştır. Diğer bir zayıflatılmış aşı,
Rotarix(Glaxo) geliştirilme aşamasındadir.
|
| |
Komplementasyon
Nükleik asit seviyesinde olmayan sadece fonksiyonel seviyede
olan etkileşim.
Örnegin, Eğer farklı genlerdeki iki ısıya duyarlı mutantı lezyonlarıyla
birlikte (ısı duyarlı) alırsak, hiçbiri yüksek sıcaklıkta (tavsiye edilmeyen)
büyüyemez. Eger aynı hücreye iki mutantı birlikte bulaştırdığımızda ise her
mutant diğerinin eksik fonksiyonlarını tamamlayabilir ve böylece
üreyebilirler (yinede viryon soyları hala mutant genomlarını icerecek ve
ısıya duyarlı olacaklardır).
Isıya duyarlı mutantları gruplamak için komplementasyonu
kullanabiliriz çünkü aynı gendeki ısıya duyarlı genler birbirini genelde
tamamlayamazlar. Bu metod mutasyonların aynı mı yoksa farklı genlerde mi
olduğunu belirlemek ve bir fonksiyonu etkileyen minimum sayıdaki geni bulmak
için genetikte kullanılan temel bir araçtır.
|
| |
Multiplisite (çesitlilik)
Reaktivasyonu Ültraviyole ışınlar kullanarak çift sarmallı DNA
virüsleri deaktive edilirse ve hücrelere cok yüksek enfeksiyon
mültiplisitisinde (mesela hücre başına bol miktarda virüs partikülleri)
deaktif virüsler bulaştırılırsa genelde reaktivasyonu görürüz. Bir viryonda
deaktive edilen genler başka birinde hala aktif olabildiği için muhtemelen
komplementasyon virüslerin baslagıçta büyümelerine izin verir. Genom
sayıları üremeden dolayi arış gösterdikçe rekombinasyon olur ve yeni
genotiplerin oluşumuna ve bazen yabani tipde virüslerin oluşumuna neden olur.
Hatalı-defektif Virüsler
Defetif virusler bulasici bir dönemi tamamlamak icin gerekli olan tam gen
tamamlanmasini yapamazlar (cogusu delesyon mutantlardir) ve bu yuzden eksik
olan fonksiyonlari tamamlamak icin baska virüslere ihtiyac duyarlar - bu
ikinci virüse yardımcı virüs denir. Defektif virüsler genomlarini
kopyalamak için ve genomlarını paketlemek için polimeraza sadece gerekli
sinyalleri sağlamak zorundadır, daha fazlasını değil. Bazı defektif virüsler
kendileri için daha fazlasını yaparlar.
Defektif virüslere örnekler: Bazı retrovirüsler konak hücresi
dizilimlerini almıştır ancak bazı viral fonksiyonlarınıda kaybetmiştir.
Bu virüsler bu fonksiyonlarıını kaybetmemiş kendilerine çok yakın
virüslere yardımcı olarak ihtiyaç duyarlar.
Bazı defektif virüsler ilgisiz virüsleri yardimcı olarak kullanırlar:
Örnegin, hepatit delta virüsü ( bir RNA virüsü) kendi zarf proteinleri
için kodlamaz ancak hepatit B virüsünün zarfını kullanır (bir DNA virüsü).
Defektif interfere edici
partiküller Yardimcı virüslerin replikasyonu eğer defektif
virüs (partikül) yoksa daha az efektif olabilir. Bunun nedeni defektif
partiküllerin yardımcı virüsün sağladiğı yardımcı fonksiyonlar için
yarışmasıdır. Bu fenomen interferans olarak bilinir ve bu fenomene sebep
olan defektif partiküller de defektif interfere edici partiküller (DI)
olarak bilinir. Tüm defektif virüsler interfere olmasada coğu öyledir.
Defektif interfere partiküllerin dogal enfeksiyonları modüle
edebileceğini not ediniz.
|
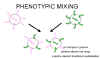 Şekil 5
Şekil 5
Aynı hücreyi enfekte eden iki farklı virüsün fenotipik karışımı |
Fenotipik karışım
Eğer iki farklı virüs bir hücreyi enfekte ederse, virüs
soyları kabuk komponentlerini iki ebeveyndende alabilir ve ebeveynlerinin
kabuk özeliklerine sahip olur. Buna fenotipik karışım denir (Şekil 5).
GENETIK MATERYALDE DEGISIKLIK OLMAZ, virüs soyları zarf doğalarına göre
değil ebeveynlerin genomlarına göre paketlenir.
Fenotipik karışım birbirine benzeyen virüsler arasında da olur örnegin
Picornavirüs ailesinin farkli üyeleri veya genetik olarak birbrinden
bağımsız virüsler arasında, örneğin Rhabdo ve Paramyxo virüsler. Bir sonraki
durumda karışan iki virüs genelde zarflanmıştır çünkü başka virüslerin
ikohedral kapsidlerinde nukleik asidi paketlemeye göre başka virüslerin
zarflarında nukleokapsitleri paketlemede daha az kısıtlama olduğu görünüyor.
|
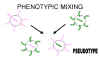 Şekil 6
Şekil 6
Pseudotip oluşturmak için fenotipik karışım |
Bazı durumlarda tamamen başka bir virüsun kabuğuyla
karşılaşılabiliriz, örneğin rhabdovirüs zarfındaki retrovirüs nükleokapsid.
Bu çeşit fenotipik karışım bazen pseudotip (pseudoviryon) formasyon olarak
tanımlanır (Şekil 6). Yukarda açıklanan pseudotip, rhabdovirüslerin
karekteristik tutunma-penetrasyon- yüzey antijenliğini gösterir ve bir
enfeksiyon anında retrovirüs olarak davranır ve retrovirüs soyları üretir.
Bunun sonuncuda pseudotiplerde değismiş konak çeşidi/dokusu tropizmi geçici
olarak gelişir.
|
|
|
 Mikrobiyoloji ve İmmünoloji On-line, Viroloji Bölümüne Dönünüz
Mikrobiyoloji ve İmmünoloji On-line, Viroloji Bölümüne Dönünüz
This page last changed on
Sunday, June 05, 2016
Page maintained by
Richard Hunt
|
 Şekil 3
Şekil 3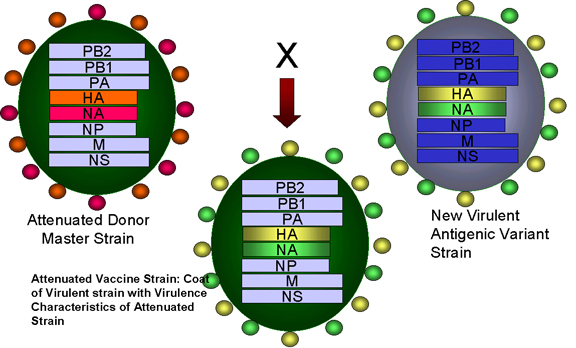 Şekil 4
Şekil 4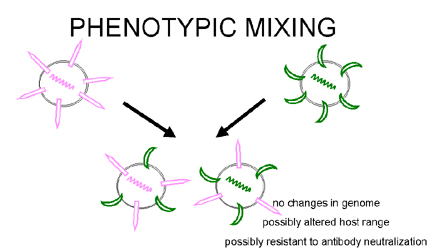 Şekil 5
Şekil 5 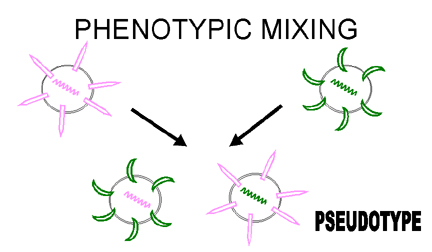 Şekil 6
Şekil 6