|
x |
x |
|
 |
 |
|
INFECTIOUS
DISEASE |
BAKTERIYOLOJİ |
İMMÜNOLOJİ |
MYCOLOGY |
PARASITOLOGY |
VİROLOJİ |
|
ENGLISH
|
VİROLOJİ - BÖLÜM DÖRT
RNA VİRÜSLERİNİN REPLİKASYON STRATEJİLERİ
Dr Margaret Hunt
Professor Emerita
Department of Pathology, Microbiology and Immunology
University of South Carolina School of Medicine
Çeviren. Prof. Dr. Selçuk Kaya
İzmir Katip Çelebi Üniversitesi, Tıp Fakültesi
|
|
Español |
|
FRANCAIS |
|
SHQIPTARE |
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|

 |
|
|
|
|
|
ÖĞRETİM HEDEFLERİ
Hayvan RNA virüsleri tarafından kullanılan replikatif stratejilerinin
tanımlayıcı analizi
Farklı RNA virüsü replikasyon şemaları ile ilişkili prototiplerin
tanımlanması
 Polio Tip 1’in yapısı Mahoney. X-ray data from Hogle et al.(Harvard
Univ.), PDB entry 2PLV, rendered with GRASP
Polio Tip 1’in yapısı Mahoney. X-ray data from Hogle et al.(Harvard
Univ.), PDB entry 2PLV, rendered with GRASP
(A.Nicholls, Columbia Univ.) Courtesy of Dr Sgro and
theInstitute for Molecular Virology, Univ. of Wisconsin (used with
permission) |
RNA VIRUS
REPLİKASYONU - GENEL
STRATEJİLER
RNA virüsleri DNA
fazı içermez
RNA’ları yoluyla replike olan virüsler çoğalabilmek için
RNA bağımlı RNA polimeraza ihtiyaç duyarlar fakat hayvan hücrelerinin
uygun enzime sahip olmadığı görülmektedir. Bu yüzden bu tip RNA
virüsleri RNA bağımlı RNA polimerazı kodlamak zorundadır.
Viral mRNA olmadan hiçbir viral protein kodlanamaz, bu
nedenle viriondaki RNAnın yapısı virüsü şu yollardan birine yönlendirir:
Pozitif Polariteli RNA
virüsleri
Bu virüslerde genomik RNA, mRNA ile aynı fonksiyon
ve aynı anlamdadır. Bu mRNA konak hücrede enfeksiyona neden
olabilmektedir.
Örnekler:
Negatif Polariteli RNA
Virüsleri
Virion RNAsı negatif polariteli yani mRNAyı
tamamlayıcıdır ve bundan dolayı proteinler sentezlenmeden önce
tamamlayıcı pozitif polariteli mRNAya kopyalanmalıdır. Bu nedenle
RNA bağımlı RNA polimeraza ihtiyaç duymanın yanı sıra, bu virüsler
hücreye enfekte edecek mRNAlarını üretebilsin diye ayrıca bu enzimin
virion içinde paketlenmesine de ihtiyaç duyarlar.
Örnekler:
-
İnfluenza virüs (orthomiksovirüs)
-
Kızamık virüsü, kabakulak virüsü (paramiksovirüs)
-
Kuduz virüsü (rabdovirüs)
Çift İplikli RNA Virüsleri
Genomik RNA çift ipliklidir ve mRNA olarak fonksiyon
göremez bu sebeple bu virüsler konak hücreyi enfekte ettikten sonra
kendi mRNAlarını yapabilmek için RNA polimerazı paketlemeye ihtiyaç
duyar.
Örnekler:
RNA’larından DNA
kopyalayabilen RNA virüsleri
Bunlar retrovirüslerdir. Bu durumda her ne kadar viral
RNA pozitif polariteli olsa da kapsitten sitoplazmaya salınıncaya kadar
mRNA olarak fonksiyon görmez. Bunun yerine, revers transkriptaz enzimi
için kalıp görevi görür ve DNAya kopyalanır. Revers transkriptaz hücrede
bulunmaz bu yüzden bu virüsler bu enzimi kodlamak ve paketlemek
durumundadır.
|
DNA FAZINA SAHİP OLMAYAN RNA
VİRÜSLERİ |
|
Genom |
RNA-bağımlı RNA polimeraz (=virionda transkriptaz) |
RNA’nın Infektivitesi |
Hücrede ilk olay |
| Pozitif polariteli RNA |
Yok |
Infektif |
Translasyon |
| Negatif polariteli RNA |
Var |
Non-infektif |
Transkripsiyon |
| Çift iplikli RNA |
Var |
Non-infektif |
Transkripsiyon |
|
RETROVIRUSLER |
|
Genom |
RNA-bağımlı RNA polimeraz (=virionda transkriptaz)
|
RNA’nın Infektivitesi |
Hücrede ilk olay |
| Pozitif polariteli RNA |
Var |
Non-infektif |
Reverse transkripsion |
TRANSLASYON SORUNU
Ökaryotik konak hücreleri translasyon sırasında protein sentezi mekanizması
olarak genellikle monosistronik mRNAlar kullanır ve bu durumda tek bir mRNAdan
birden fazla tip protein sentezinde problem oluşur.
RNA virüslerinin bu sorunla ilgili çeşitli çözümleri vardır:
- Virüs birçok monosistronik mRNA yapar
- Virüs birden çok monosistronik RNA oluşturabilmek için konak splicing
mekanizmasıyla işlenmiş primer transkriptleri yapar
- Viral mRNA monosistronik transkript gibi davranır. Sonradan parçalara
bölünecek olan poliprotein olarak adlandırılan büyük bir polipeptid yapılır.
Böylece bu ilk translasyonu ürününden birden çok protein işlenmiş olur. Örn:
picornavirüsler
- Viral mRNA, ribozomları 5’ucu yerine içerden bağlayabilmeyi sağlayan
özel özelliklere sahiptir.
RNA VİRÜSLERİNDE GENOM BÜYÜKLÜĞÜ
RNA virüsleri göreceli olarak küçük genoma sahip olma eğilimindedir (her
ne kadar virion boyutu küçük olmak zorunda olmasa da). Bu büyük ihtimalle
RNA tamir mekanizmalarının genom büyüklüğü üzerine koyduğu sınırlamalardan
kaynaklanmaktadır.
Küçük bir genoma sahip olmanın sonucu RNA virüsleri yalnızca birkaç
protein kodlama eğilimindedir. Viral bağlantı proteinleri ve RNAdan
tamamlayıcı nükleik asit (RNA veya retrovirüsler için DNA) kodlayan bir
polimerazı içermektedir.
|
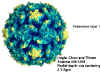 Şekil 1
Şekil 1
Polio virus
© J-Y Sgro, Used with permission. From Virus World
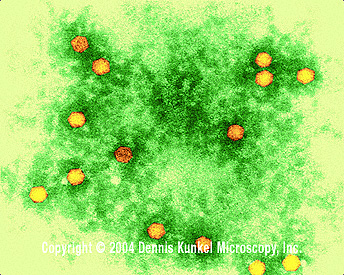
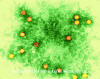 Şekil 2
Şekil 2
Polio virus x350,000
© Dennis Kunkel Microscopy, Inc. Used with permission |
POZİTİF
POLARİTELİ RNA VİRÜSLERİ
Örnekler:
-
Picornavirüsler
-
Togavirüsler
-
Flavivirüsler
PİCORNAVİRÜSLER (PİCORNAVİRİDEA)
Özellikler
Küçük (28nm), çıplak ikozahedral simetrili
virüslerdir(pico: çok küçük). RNAları tek iplikli, pozitif polariteli,
poliadenillidir. Enfeksiyon durumunda doğrudan mRNA olarak görev yapar.
Prototipi: poliovirüs (şekil 1, 2)
|
 Şekil 3
Şekil 3
Picornaviridae genomik RNA’nın yapısı
|
Tutunma ve
Penetrasyon
Bir viral protein konak hücre membranında reseptörünü
ayırd eder (bu viral tropizm açısından önemlidir). Görünen o ki
reseptöre bağlanmak kapsit yapısını bir şekilde değiştirir, hücre
membranında bir kanal oluşur ve RNA sitoplazmaya salınır. Artık mRNA
translasyon için hazırdır.
|
 Şekil 4
Şekil 4
Adapted from Schaechter et al., Mechanisms of Microbial
Disease, 2nd Ed. |
Viral Protein
Sentezi
Poliovirüsün viral RNAsı mRNA gibi fonksiyon görmektedir
fakat tipik ökaryotik mRNAları gibi metillenmiş başlık yapısı
içermemektedir- 5’metillenmiş başlık yapısını tanıma zorunluluğu olmadan
ribozomlara tutunabilmeyi sağlayan ribozom bağlantı noktalarına sahiptir
(içerden ribozoma giriş bölgesi veya IRES) (Şekil 3).
Picornavirüsler sıklıkla konak hücrede metillenmiş
başlık tanımasını engellerler. Pekçok konak hücrede translasyon başlık
bağımlıdır, yani bu durum konak protein sentezinin birçoğunu inhibe
ederken viral protein sentezini etmez- tek yolu bu virüslerin konak
hücreyi kendi faydası yönünde değiştirmesidir.
mRNA kırpılmış tek bir polipeptide (poliprotein)
dönüştürülür. Bu kırpılma translasyon tamamlanmadan evvel oluşur(örn
büyüyen zincir) ve virüs tarafından kodlanan proteazlarla yapılır (şekil
4). Bu proteazlardan bazıları poliproteinin parçaları olsa dahi
çalışabilirler.
Kırpılma sonucu oluşan ürünler:
|
 Şekil 5
Şekil 5
Picornaviridae viral genomun Replikasyonu
|
RNA Replikasyonu
Replikasyonu sürdürmek için yeni viral protein yapımına
ihtiyacımız var.
1. Viral RNA polimeraz pozitif polariteli genomik RNA’yı
tamamlayıcısı negatif polariteli RNA ya kopyalar:
Bu işlem
-
VPg, (veya prekürsör içeren VPg)
-
Viral RNA polimeraz (replikaz) ve
-
Konak hücre proteinlerine ihtiyaç duyar.
VPg RNA sentezinde primer gibi davranır, bu tüm yeni
sentezlenmiş RNA moleküllerinin 5’ ucunda bulunmasını açıklar.
2. Yeni negatif polariteli iplik, yeni pozitif polariteli
iplik için kalıp görevi görür (Şekil 5). Poliovirüs RNA polimeraz ve VPg ye
ihtiyaç vardır. VPg yeni pozitif polariteli zincirin 5’ ucuna bağlıdır (büyük
ihtimalle primer olarak fonksiyon görür).
Yeni pozitif polariteli ipliğin üç alternatif yolu vardır:
-
Diğer negatif polariteli zincirlere kalıp olabilir
-
Yeni oluşan virionlar içinde paketlenebilir
-
Poliproteinlere dönüşebilir (bu durumda VPg
translasyondan önce kaldırılır)
Birleşme
Yeterli derecede pozitif polariteli RNA ve viral proteinler
biriktiği zaman toplarlanma başlar. Partiküller içerde VPg-RNA ve kapsitte
üç tip protein (VP0, 1 ve 3) olmak üzere toplanır. Virion mature hale
geldeğinde VP0 proteini VP2 ve VP4e parçalanır, ve bu virion enfektiftir.
Takip eden hücre lizisi ile virionlar salınır. Fazlalık kapsitler şekillenir
ve inklüzyon cisimcikleri sitoplazmada görülebilir.
NOT: TÜM YAŞAM DÖNGÜSÜ SİTOPLAZMADA GERÇEKLEŞİR
ERKEN VE GEÇ GEN EKSPRESYONU OLARAK AYRIM YOKTUR
|
 Şekil 6
Şekil 6
Balık Epitel hücresinde Rhabdovirus
© Dennis Kunkel Microscopy, Inc. Used with permission |
NON-SEGMENTLİ
NEGATİF ZİNCİRLİ VIRUSLAR
Segmentsiz negatif zincirli RNA viruslarına Örnekler:
-
Rabdovirüsler (Şekil 6). Rabies virus, vesicular
stomatit virus, Mokola virus, Duvenhage virüsü içerir
-
Paramiksovirüsler (Şekil 11). Newcastle disease virus,
parainfluenza viruses,mumps virus, measles virus, respiratory syncytial
virusu içerir.
|
 Şekil 7
Şekil 7
Tipik rhabdovirus yapısı
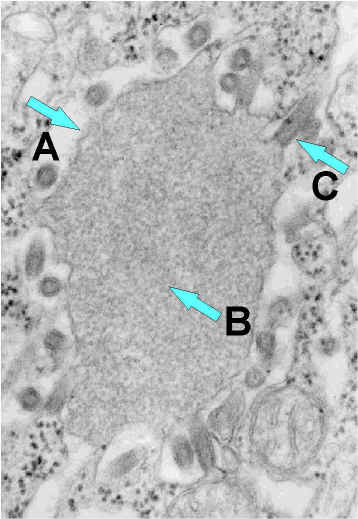
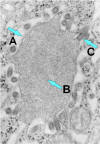 Şekil 7b
Şekil 7b
Rabies virus sinir hücresi endoplazmik retikulumunda inkluzyon (Negri
cisimciği) A. Negri cisimciği. B. dikkat inklüzyonda bol RNA. C. rabies
virüs tomurcuklanması.
CDC
|
RHABDOVIRUSLAR (RHABDOVIRIDAE)
Örnek: Rabies virus. En yoğun çalışılan üyesi vesicular stomatitis
virus. RNA genom:
- Tek zincirli
- Negatif (minus) polariteli
- 5 protein kodlar
Tutunma, Penetrasyon ve
Soyunma
Virüs hücre yüzeyinden absorbe edilir.
G (glikoprotein) konak hücre yüzeyinde reseptöre bağlanan
bağlantı proteinidir (Şekil 7). Bağlanan virüs endositozla içeri
alınır. Viral membran ile endozom membranı füzyona uğrar (endozomun
asit ph sı önemlidir çünkü füzyondan önce G proteinler asit ph ya
maruz kalmalıdır). Viral membran ile endozom membranının füzyonu
sonucunda nükleokapsit sitoplazmaya salınır.
Transkripsiyon
Bu metinde ‘transkripsiyon’ mRNA sentezine işaret etmektedir.
Transkripsiyon için nükleokapsitten tam bir soyunma gerekli değildir-
nükleokapsit formda bulunurken viral RNA polimeraz viral RNAyı
kodlayabilir. Genomik RNA bir dereceye kadar ribonükleazlardan
korunduğu için bu bir avantajdır.
Kodlanan her beş viral protein için bir monosistronik mRNA
bulunmaktadır. Bu mRNA lar başlıklı, metillenmiş ve
poliadenillenmiştir. Negatif polariteli RNA virüsü, sitoplazmik
olduğu için, mRNA sentezi ve modifikasyonu yapan enzimler virionda
paketlenmiş halde hazır bulunur.
Translasyon
Haberci RNAlar konakçı ribozomlarında translasyona uğrar ve beş
viral protein de aynı anda üretilir. Erken ve geç fonksiyonlarda
herhangi bir fark yoktur.
|
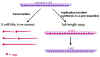 Şekil 8
Şekil 8
Rhabdovirus RNA’nın replikasyon ve transkripsiyonu
|
RNA replikasyonu
RNA replikasyonu genom uzunluğunda yeni RNA kopyalarının
yapıldığı bir prosedürdür (Şekil 8). RNA replikasyonu sitoplazmada
gerçekleşir ve viral RNA polimeraz tarafından gerçekleştirilir. Pozitif
zincirin tamamı sentezlenir sentezlenmez nükleokapsit proteinleriyle
kaplanır (mRNA, konak proteinlerinin translasyonuna engel olan bu
proteinlerle kaplı değildir).
Yeni pozitif zincir, nükleokapsit proteinleriyle kaplı
negatif zincirden kopyalanır. (not: viral RNA polimeraz mRNA (transkripsiyon)
ve RNA sentezler (replikasyon ), transkriptaz veya replikaz olarak da
adlandırılır, bu adlandırma ile polimeraz aktivitesinin farklı yönlerine
dikkat çekmektedir.)
Yeni negatif zincir:
|
 Şekil 9
Şekil 9
Endoplazmik retikulumdan plazma membranına glikoprotein transferi
|
Birleşme
Virüs iki modül içerir- zarf ve nükleokapsit:
Zarf
Endoplazmik retikuluma bağlı ribozomlarda transmembran proteinler
yapılır. Yapıldığı gibi endoplazmik retikulum membranına
yerleştirilir, endoplazmik retikulumda glikozillenir ve karbonhidrat
zincir modifikasyonları için golgi cisimciğine aktarılır. Ardından
veziküller içinde hücre membranına transporte edilir, veziküler
stomati virüsü için bu plazma membranıdır (Şekil 9).
|
 Şekil 10
Şekil 10
Rhabdovirus birleşmesi
|
Nükleokapsit
Nükleokapsit sentezi yukarıda tanımlandı. Viral RNA polimeraz
kompleksi yapılır yapılmaz nükleokapsitlerle bağlantı haline geçer.
Nükleokapsit, G ve M protein içeren membranın modifiye alanlarına
doğru tomurcuklanır. M (Matrix ) proteini toplanma aşamasında yer
alır.- G proteinin memranda bulunan parçalarıyla ve
nükleokapsitlerle etkileşim halindedir.
NOT:
Tüm yaşam döngüsü sitoplazmada gerçekleşir.
RNA polimeraz ve RNA modifikasyon enzimleri virüs tarafından
kodlanır ve virion içinde bulunur.
Erken / geç bölünme yoktur.
|
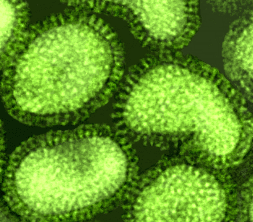
 Şekil 11 Paramyxovirus
Şekil 11 Paramyxovirus
© Dr Linda Stannard,University of Cape Town, South Africa
(used with permission) |
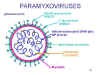
PARAMİKSOVIRUSLAR (PARAMYXOVIRIDAE)
Paramiksovirüsler pleomorfiktir (Şekil 11), yani bir popülasyonda
virüsün birden fazla morfolojik formu vardır. Negatif iplikli,
segmentsiz RNA ve helikal nükleokapsitleri vardır (Şekil 12). Konak
hücre membranı kaynaklı bir zarfla çevrilidirler.
Zarf virüs tarafından kodlanan iki glikoprotein içerir: F protein
ve bağlantı proteini
- F proteinin füzyon fonksiyonu vardır
- Bağlantı proteini konak hücrede reseptöre bağlanır
Bu protein hemaglütinin ve nöraminidaz aktivitesinin ikisine
birden (HN protein) veya yalnızca hemaglütinin (H protein) ya da
(G) aktivitesine sahip olabilir.
|
 Şekil
12
Şekil
12
Tipik paramyxovirus yapısı |
|
PARAMYXOVIRUS AİLESİ YÜZEY
GLİKOPROTEINLERİ |
|
GENUS
|
GLİKOPROTEIN
|
TiPİK ÜYE
|
|
PARAMYXOVIRUS AİLESİ |
|
Paramyxovirus
|
HN, F
|
HPIV 1
HPIV 3
|
|
Rubulavirus |
HN, F |
HPIV 2
HPIV 4
mumps virus |
|
Morbillivirus
|
H, F
|
measles virus
|
|
PNEUMOVIRUS AİLESİ |
|
Pneumovirus
|
G, F
|
respiratory syncytial
virus
|
|
Metapneumovirus |
G, F |
metapneumoviruses |
Hemaglütinasyon
Hemaglütinasyon, klinik laboratuvarlarda kolaylıkla test
edilebilir ve tanı amaçlı kullanılmaktadır.
Hemaglütinasyon eritrositlerin hemaglütine olmasını ve virüsün
eritrositlerdeki reseptörlere bağlanabilmesini sağlamaktadır.
Her bir virion birden fazla bağlantı proteini içerdiğinden,
birden fazla eritrosite bağlanabilir ve böylece onların bir ağ
halinde bağlanmasını sağlar. İnaktive virüs, bağlantı
proteinleri sağlam olduğu sürece hemaglütinasyon yapabilir.
Eğer bir kişide viral hemaglütinine karşı antikor bulunuyorsa,
bu antikorlar bağlantı proteinlerine bağlanır ve eritrositlere
bağlanmasını engeller. Antikor içerdiğinden dolayı bu kişinin
serumunun aglütine olması önlenmiş olur- fakat diğer hemaglütine
edebilen virüsler için geçerli değildir. Bu bir kişinin hangi
hemaglütine edici virüsle enfekte olduğunu saptamak için
kullanılabilir.
Hemadsorbsiyon
Enfeksiyon süresince, viral bağlantı proteinleri enfekte
hücrenin plazma membranına yerleştirilir. Eğer viral bağlantı
proteinleri eritrositlere bağlanabilirse, enfekte hücre
eritrositlere bağlanabilir çünkü yüzeyinde bağlantı proteini
içermektedir- bu hemadsorbsiyon olarak adlandırılır. Klinik
laboratuvarlarda bu durum enfeksiyonun erken evrelerinde virüsle
enfekte hücrelerin tanınmasına ve hücrelere gözle görülebilir
bir hasar vermeden tanınmasına olanak sağlar.
|
 Şekil 13
Şekil 13
Paramyxovirusların endositozu ve tutunması
|
Tutunma ve
penetrasyon
H(N)/G protein hücre yüzeyindeki reseptörleri tanır.
F proteini fizyolojik pHda membranlar arası füzyonu
kolaylaştırır, her ne kadar paramiksovirüsler endositozla alınsa da,
plazma membranından direkt füzyon ile de hücreye girebilirler (Şekil
13).
F proteininin fizyolojik pHdaki fonksiyonları sayesinde,
paramiksovirüs enfeksiyonları sinsitya oluşumuyla sonuçlanır (bkz-
fizyolojik pHda füzyonun sonuçları, DNA virüs replikasyon
stratejileri-herpesvirüs).
|
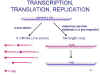 Şekil 14
Şekil 14
Paramyxovirus RNA’nın replikasyon ve transkripsiyonu
|
Transkripsiyon,
translasyon ve RNA’nın replikasyonu
Hücre içi olaylar rhabdovirüs gibidir (Şekil 14):
-
Viral çoğalma sitoplazmadadır
-
Viral RNA polimeraz kalıp olarak nükleokapsiti
kullanır
-
Viral RNA polimeraz çıplak bir nükleokapsite
gereksinim göstermez
-
Viral mRNA transkripsiyona uğrar, başlığı takılmış,
metillenmiş ve poliadenillenmiştir
-
Negatif polariteli RNA virüsü olduğundan dolayı RNA
polimeraz ve RNA modifikasyon enzimleri virionda paketlenmiştir
-
Viral mRNA lar protein üretmek üzere translasyona
uğrar
-
Gen ekspresyonunda erken ve geç fonksiyonlar
açısından bir fark yoktur
Viral RNA replikasyonu pozitif zincir üretimini içerir. Bu negatif
zincir için kalıp görevi görür. Her iki zincir de yapılır yapılmaz
nükleokapsit proteinleriyle kaplanır (Şekil 14).
Yeni negatif zincir replikasyon veya transkripsiyon için
kalıp görevi görür veyahut yeni virionlarda paketlenir.
|
 Şekil 15
Şekil 15
Proteolitik ayrılma ile fuzyon proteinin aktivasyoun
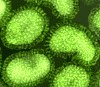 Şekil 16 Orthomyxovirus (Influenza A)
Şekil 16 Orthomyxovirus (Influenza A)
© Dr Linda Stannard, University of Cape Town, South
Africa
|
Birleşme
Viral glikoproteinlerin her ikisi de (bağlantı proteini
ve F füzyon proteini) transmembran proteinlerine dönüştürülür ve plazma
membranına gönderilir. M (matriks) proteini, glikoproteinlerin
yerleştirildiği plazma membranı bölgeleri ile nükleokapsitlerin
etkileşimini sağlar.
Virüs membrandan tomurcuklanarak ayrılır.
Nöraminidazın
Fonksiyonu
Paramiksovirüslerde nöraminidaza sahip olan suşların
hücreden salınımı daha kolaydır. Bu virüslerde reseptörün önemli bir
parçası olarak sialik asit bulunmaktadır. Nöraminidaz hücre yüzeyinden
sialik asiti (nöraminik asit) uzaklaştırır. Böylece, sialik asit hücre
yüzeyinden uzaklaştırıldığında ve progeny virionlar fonksiyonel
reseptörlere sahip olmadığından, progeny virüsler bir diğerinden veya
tomurcuklandığı hücreden veya herhangi bir enfekte hücreden ayrılabilir.
Böylelikle henüz enfekte olmamış hücrelere doğru yayılabilirler.
Nöraminidaz enfeksiyonun yayılmasına yardım eder, eğer
virüs mukus içinde sialik asit rezidülerine bağlanırsa, reseptörüne
beğlanamaz ve o hücreyi enfekte edemez. Fakat eğer mukustaki sialik asit
hasarlanmışsa, virüs salınabilir ve hücre yüzeyindeki reseptörlerine
ulaşabilir.
F Protein
Aktivasyonu
Virüs başka bir hücreye bağlandığında füzyonu aktive
edebilmek için F proteini kırpılmalıdır (Şekil 15). Bu maturasyonun geç
olaylarındandır.
|
Rhabdovirus ve
paramyxovirus arasındaki bazı farklar |
| |
Rhabdovirus |
Paramyxovirus |
| Biçim |
Mermi
Basiliform |
Yuvarlak
Pleomorfik |
| Glikoprotein |
Bir (hem tutunma hem de füzyon) |
İki (bir tutunma ve bir fuzyon) |
| Füzyon pH |
Asidik |
Nötral
Fizyolojik |
|
|
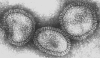 Şekil 17 Orthomyxovirus (Influenza A) © Dr Linda Stannard,
University of Cape Town, South Africa
Şekil 17 Orthomyxovirus (Influenza A) © Dr Linda Stannard,
University of Cape Town, South Africa
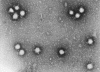 Şekil 18 Bunyavirus
Şekil 18 Bunyavirus
From ICTV database
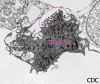 Şekil 19b Vero E6 doku kültür hücrelerinin arenavirus ile infekte edilmesi.
Resim hücre yüzeyinden ekstrasellular virus partiküllerinin tomucuklanmasını
gösteriyor. Büyütme yaklaşık. 12,000 kez. Image courtesy
Cynthia Goldsmith, MS, Infectious Disease Pathology Activity, DVRD, NCID,
CDC
Şekil 19b Vero E6 doku kültür hücrelerinin arenavirus ile infekte edilmesi.
Resim hücre yüzeyinden ekstrasellular virus partiküllerinin tomucuklanmasını
gösteriyor. Büyütme yaklaşık. 12,000 kez. Image courtesy
Cynthia Goldsmith, MS, Infectious Disease Pathology Activity, DVRD, NCID,
CDC |
SEGMENTLİ NEGATİF ZİNCİRLİ
VIRUSLAR Örnekler:
- Orthomyxoviruslar (Şekil 16, 17)
- Bunyaviruslar (Hantavirus genusu dahil) (şekil 18)
- Arenaviruslar (Şekil 19b)
ORTHOMİKSOVİRUSLAR (ORTHOMYXOVIRIDAE)
Üç grup influenza virüsü vardır: A, B ve C. İnfluenza A üzerinde en
çok çalışılan olup, influenza A ve B insanlar için en önemlileridir.
İnfluenza virüsleri pleomorfiktir (yani farklı şekillerdedir).
Negatif polariteli, tek zincirli RNA ve segmentli RNA genomuna
sahiptirler. İnfluenza A da sekiz adet segment bulunur. Nükleokapsitleri
helikaldir (Şekil 19). RNA polimerazı virionda paketlenmiştir.
Bu virüsler zarflıdır ve iki membran glikoproteini içerir (Şekil 19):
- HA-Hemaglütinin- bağlantı ve füzyon proteini
- NA-Nöraminidaz- salınımda önemlidir. Konak hücre ve viral
proteinlerden sialik asiti ayırır.
|
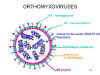 Şekil 19
Şekil 19
Tipik orthomyxovirus yapısı |
Adsorbsiyon ve Penetrasyon
Virüs hücre yüzeyindeki reseptörlere bağlanır ve endositoz ile içeri
alınır. Endozomun asit pHsında HA konformasyonel değişim gösterir ve
füzyon oluşur. Nükleokapsit sitoplazmaya salınır.
|
 Şekil 20
Şekil 20
Orthomyxoviridae RNA’nın transkripsiyonu
|
Transkripsiyon, Translasyon ve
Replikasyon
Nükleokapsit nükleusa taşınır. mRNA sentezi ve viral RNA replikasyonu
nükleusta gerçekleşir. Bu bir RNA virüsü için alışılmadık bir durumdur.
İnfluenza virüsünün mRNAsının metillenme ve 5’ ucuna başlık takılma
işlemi de farklı mekanizmalarla olmaktadır.
İnfluenza virüsünde paketlenmiş olan viral endonükleaz 5’ ucundaki
başlığı, metillenmiş mRNA’yı 5’ ucundan yaklaşık 13-15 bazlık kısmını
keser ve bunu viral mRNA sentezi için primer olarak kullanır (Şekil 20)-
bundan dolayı konak mRNAsından kaynaklanan tüm influenza mRNA ları 5’
ucunda kısa bir uzunluğa sahiptir.
Viral RNA polimeraz (transkriptaz) primeri devam ettirir ve
tamamlayıcı pozitif zincirli mRNAyı kalıp olarak kullanıp kopyalar ve
poli(A) kuyruğu ekler. Transkripsiyon, her segmentte bir transkript
olmak üzere 8 adet transkriptle sonuçlanır. Bazı segmentler alternatif
kırpılma ile primer transkriptlerini uzatır (influenza virüsünün RNA
sentezi nükleusta gereçekleştiğinden, kırpılma işlemine tabi tutulabilir),
her biri iki adet alternatif transkript oluşturabilir. Örneğin M
segmenti iki alternatif mRNA oluşturabilir. Bunlar M1 ve M2 protein
olarak kodlanır. Böylelikle virüs kırpılma mekanizmasına sahip olduğu
sürece tek bir segment birden fazla proteine kodlanabilir. mRNAlar
sitoplazmada translasyona uğrar. RNA replikasyonu için gereken
proteinler nükleusa gönderilirken transmembran proteinler plazma
membranına hareket eder.
|
| |
RNA Replikasyonu
RNA replikasyonu virüsün kodladığı bir enzimle nükleusta gerçekleşir
(mRNA transkripsiyonundaki RNA polimerazla aynı veya modifiye bir
versiyonu). Virion RNAsının aynı uzunlukta, tam bir tamamlayıcı kopyası
yapılır- bu pozitif polariteli RNA yapılır yapılmaz nükleokapsit
proteinleriyle kaplanır. Bu pozitif zincir RNA daha sonra negatif zincir
sentezi için kalıp olarak kullanılır; tekrardan yeni negatif zincir
yapıldığı gibi nükleokapsit proteinleriyle kaplanır. Yeni negatif zincir
replikasyon ve mRNA sentezi için kalıp olarak kullanılabilir veya
paketlenir.
Birleşme
Plazma membranında gerçekleşir. Zarf proteinleri golgi cisimciğinden
plazma membranına gönderilirken nükleokapsit proteinleri nükleus dışına
gönderilir. M1 proteini hem nükleokapsitle hem de HA ve NA
glikoproteinlerini içeren plazma membranının modifiye bölgesiyle
bağlantı içindedir. Bundan sonra virüs konak hücre membranına doğru
tomurcuklanır.
NOT:
- HA füzyondan önce kırpılmalıdır. Kırpılma virüs hücreden
ayrılınca veya ekstrasellüler sıvıda gerçekleşir. Kırpılma hangi
dokunun infektif virüs üretebileceğinin belirleyicisidir. Füzyona
neden olmadan önce bir sonraki hücreyi enfekte ettiklerinde
genellikle asidik endozom ortamına maruz kalmaları sebebiyle
kırpılmış proteinlerin konformasyonel değişim geçirmeleri gerekir.
- NA, reseptörlerden sialik asiti uzaklaştırıp virüsün hücreden
ayrılmasına yardım eder. NA ayrıca virüsün respiratuar sistemdeki
epitel hücrelerine ulaşması için mukus içindeki sialik asit
reseptörlerinden ayrılmasını sağlayarak mukusa tutunmasına, yardımcı
olur. Nöraminidaz virüsün yeni hücreleri enfekte etmesini önlemez
çünkü muhtemelen endositoz, reseptör ayrılmasından daha hızlıdır.
Paramiksovirüs ailesi ile Orthomiksovirüs ailesi arasında
benzerlikler ve farklar bulunmaktadır, her iki ailenin üyeleri de
zarflıdır, her ikisi de negatif zincirlidir, tek zincirli RNA ve
helikal nükleokapsit içerirler. Fakat her iki aile birbirinden çok
farklıdır. İki aile arasında immünolojik ilişki bulunmamaktadır.
|
| |
|
ÖZELLİK
|
PARAMYXOVIRIDAE
|
ORTHOMYXOVIRIDAE
|
|
Genom
|
Segmentsiz
|
Segmentli
|
|
RNA sentezi
|
Sitoplazmik
|
Nükleer
|
|
mRNA primer ihtiyacı
|
Hayır
|
Evet
|
|
Hemaglutinin,nöraminidaz
|
Eğer ikisi varsa, aynı protein kısmı (HN)
|
Influenza A ve B ikisini de fakat 2 farklı protein (HA
ve NA)
|
|
Sinsitya formasyonu
|
Evet (F fonksiyon normal pH’da)
|
Hayır (HA fonksyion asid pH’da)
|
|
 Şekil 21 Memeli Reovirus Virion
Şekil 21 Memeli Reovirus Virion
The cryoEM data was from Tim Baker's Laborratory, Purdue University. Movies
were created by Stephan Spencer.
Copyright 1999 Dr Tim Baker and Stepthen M Spencer. From Dr
J-Y Sgro's Virusworld
|
ÇİFT ZİNCİRLİ RNA
VIRUSLARI
REOVIRUS AİLESİ (REOVIRIDAE)
Reovirus ailesi:
- Reovirüs cinsinin üyeleri
- Rotavirüs cinsinin üyeleri
- Orbivirüs cinsinin üyeleri (ör. Bluetongue virus)
- Colorado tick ateşi virüsünü içerir (ör. Colorado tick fever
virus).
|
 Şekil 22 Tipik reovirus yapısı
Şekil 22 Tipik reovirus yapısı
Adapted from Joklik et al. Zinsser Microbiology 20th Ed. |
Reovirüsler ikozahedral simetri ve çok katmanlı kapsit (iç ve dış kapsit)
bulundururlar (Şekil 22). RNAsı çift zincirlidir. 10-12 segmentlidir (hangi
reovirüs familyasından hangi cins olduğuna bağlı olarak) (Şekil 22).
Reovirüs ve rotavirüs ailesi üyelerinin yaşam döngüsünde bazı önemli
farklılıklar vardır. İnsanlardaki klinik öneminden dolayı rotavirüslere
odaklanmalıyız.
|
 Şekil 23 Rotavirus (çift-kapsid partikülü (sol), ve tek, iç kapsid (sağ))
Şekil 23 Rotavirus (çift-kapsid partikülü (sol), ve tek, iç kapsid (sağ))
Copyright Dr Linda Stannard, University of Cape Town, South
Africa |
ROTAVIRUSLAR
(rota = tekerlek (elektron
mikroskoptaki görünümlerinden dolayı)(Şekil 23)
Tutunma,
Penetrasyon ve Soyunma
Rotavirüsün hücreye girişi
sırasında in vivo ortamda tam olarak ne olduğu net değildir. Kapsitin
dış yüzeyinin bazı alanlarının kesilmesi için bir proteaza ve virüs
sitoplazmaya girmeden önce intermediate subviral partikül (ISVP)
oluşturmaya ihtiyacı vardır. İn vivo olarak gastrointestinal sistemde
intermediate subviral partiküller proteaz sindirimi ile
oluşturulmaktadır. Ardından intermediate subviral partikülde büyük
ihtimalle uç noktada viral bağlantı proteini açığa çıkar ve konak
reseptörlerine bağlanır. Aktive olmuş intermediate subviral partikül
direkt olarak veya endositoz yoluyla sitoplazmaya girer. Sitoplazmada
virion RNAsı, viral RNA polimeraz tarafından kopyalanr. Bu esnada hala
daha nükleokapsit içindedir. Bu nükleokapsitin ISVP veya virionla
ilişkili olandan daha az kendisiyle ilişkili proteini bulunmaktadır.
Transkripsiyon ve
translasyon
Çift zincirli RNA, mRNA gibi fonksiyon göremez bu yüzden ilk işlem mRNA
üretmektir (transkripsiyon). mRNAlar virüs tarafından kodlanan, virionda
paketlenmiş RNA polimeraz tarafından üretilir. RNA virionda paketlenmiş
enzimlerce metillenir ve başlık takılır. Ardından kapsitin köşesine
doğru gönderilir.
mRNA translasyona uğrar ve
immatür kapsiti oluşturmak üzere viral proteinler bir araya gelir. mRNA
immatür kapsitte depolanır ve çift zincirli RNA oluşturmak için kapsit
içinde kopyalanır (virüsün her bir partikül için 11 farklı mRNAdan tek
bir kopya oluşturmasını nasıl sağladığı bilinmemektedir) (Şekil 24).
Yeni şekillenmiş immatür kapsitten daha fazla mRNA üretilebilir.
|
 Şekil 24
Şekil 24
Reoviridae’nın replikasyonu
|
Birleşme
Daha fazla protein üretilir ve en sonunda immatür kapsitler
endoplazmik retikulum lümeninden tomurcuklanır. Bu şekilde yaparak,
matür hale geldiklerinde kaybedecekleri geçici bir zarf oluşturmuş
olurlar. Bu rotavirüslerin oldukça garip bir özelliğidir.
Salınma
Büyük ihtimalle hücre lizisi ile olur.
NOT: Tüm replikasyon siklusu sitoplazmada gerçekleşir.
|
|
 Mikrobiyoloji ve İmmünoloji On-line, Viroloji Bölümüne Dönünüz
Mikrobiyoloji ve İmmünoloji On-line, Viroloji Bölümüne Dönünüz
This page last changed on
Tuesday, May 31, 2016
Page maintained by
Richard Hunt
|
 Şekil 1
Şekil 1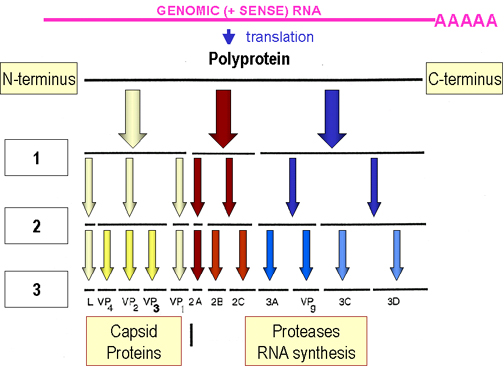 Şekil 4
Şekil 4 Şekil 6
Şekil 6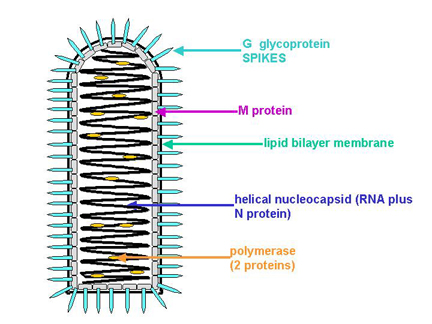 Şekil 7
Şekil 7 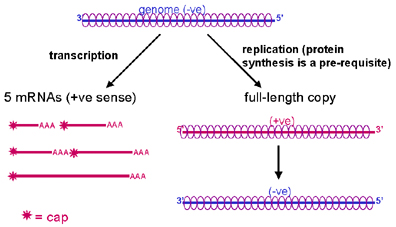 Şekil 8
Şekil 8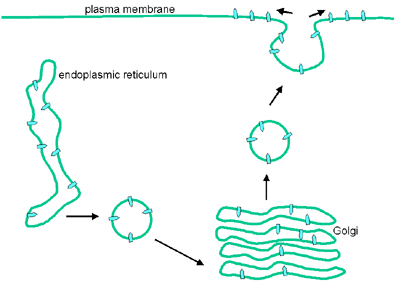 Şekil 9
Şekil 9  Şekil 10
Şekil 10  Şekil 11 Paramyxovirus
Şekil 11 Paramyxovirus  Şekil
12
Şekil
12 Şekil 13
Şekil 13 Şekil 14
Şekil 14 Şekil 15
Şekil 15 Şekil 19
Şekil 19 Şekil 20
Şekil 20 Şekil 21 Memeli Reovirus Virion
Şekil 21 Memeli Reovirus Virion  Şekil 22 Tipik reovirus yapısı
Şekil 22 Tipik reovirus yapısı  Şekil 23 Rotavirus (çift-kapsid partikülü (sol), ve tek, iç kapsid (sağ))
Şekil 23 Rotavirus (çift-kapsid partikülü (sol), ve tek, iç kapsid (sağ))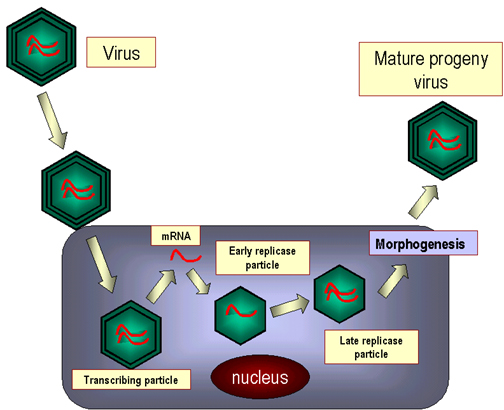 Şekil 24
Şekil 24