| x | x | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BACTERIOLOGY | IMMUNOLOGY | MYCOLOGY | PARASITOLOGY | VIROLOGY | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Dr Abdul
Ghaffar |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FRANCAIS | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| SHQIP | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PORTUGUES | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Let us know what you think |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Logo image © Jeffrey Nelson, Rush University, Chicago, Illinois and The MicrobeLibrary | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
TEACHING OBJECTIVES Know
the MHC loci and their products
|
DEFINITIONS
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
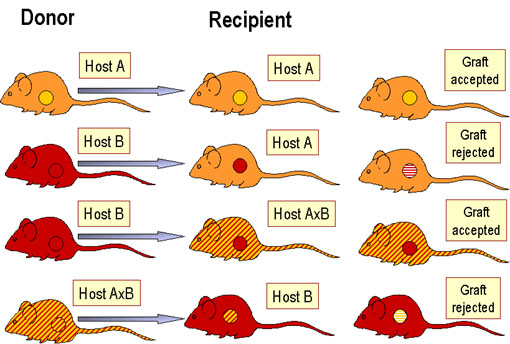 Figure 2 Laws of transplantation
Figure 2 Laws of transplantation
|
PRINCIPLES OF TRANSPLANTATION (figure 2) An immunocompetent host recognizes the foreign antigens on grafted tissues (or cells) and mounts an immune response which results in rejection. On the other hand, if an immunocompromised host is grafted with foreign immunocompetent lymphoid cells, the immunoreactive T-cells in the graft recognize the foreign antigens on the host tissue, leading to damage of the host tissue.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
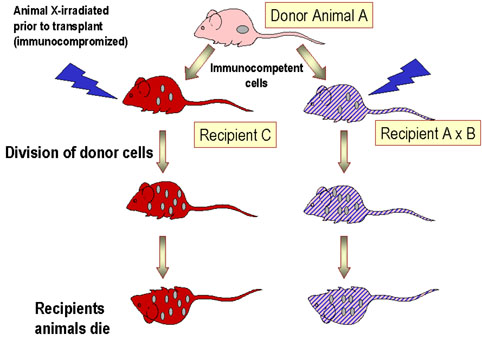 Figure 3 Graft versus host disease
Figure 3 Graft versus host disease |
Histocompatible lymphoid cells, when injected into an immunocompromised host, are readily accepted. However, the immunocompetent T lymphocytes among the grafted cells recognize the alloantigens and, in response, they proliferate and progressively cause damage to the host tissues and cells. This condition is known as graft-versus-host (GVH) disease (figure 3) and is often fatal. Common manifestations (figure 4) of GVH reaction are diarrhea, erythema, weight loss, malaise, fever, joint pains, etc. and ultimately death.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
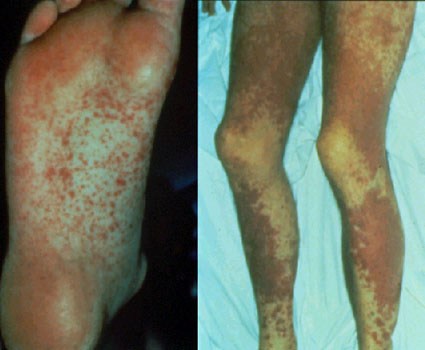 Figure 4 Graft versus host disease
Figure 4 Graft versus host disease |
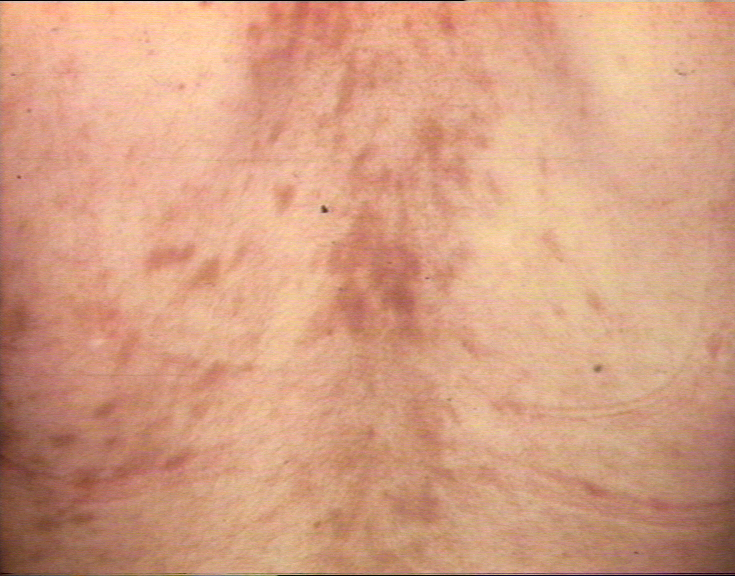
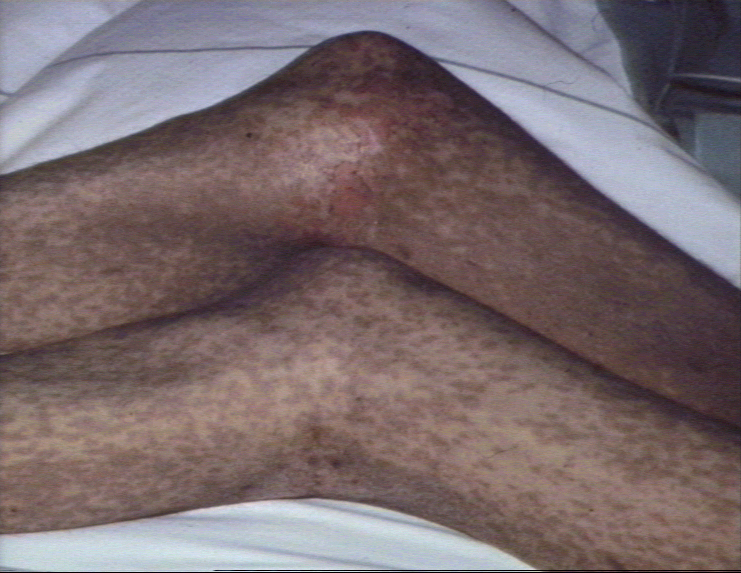 Early, chronic graft-versus-host reaction with widespread, almost confluent hyperpigmented lichenoid papules and toxic epidermal necrosis-like appearance on knee
© Bristol Biomedical Image Archive. Used with
permission
Early, chronic graft-versus-host reaction with widespread, almost confluent hyperpigmented lichenoid papules and toxic epidermal necrosis-like appearance on knee
© Bristol Biomedical Image Archive. Used with
permission
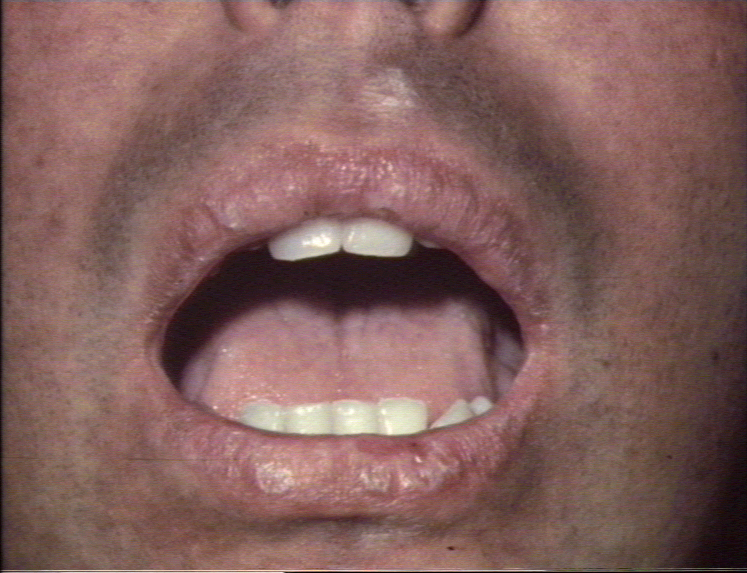
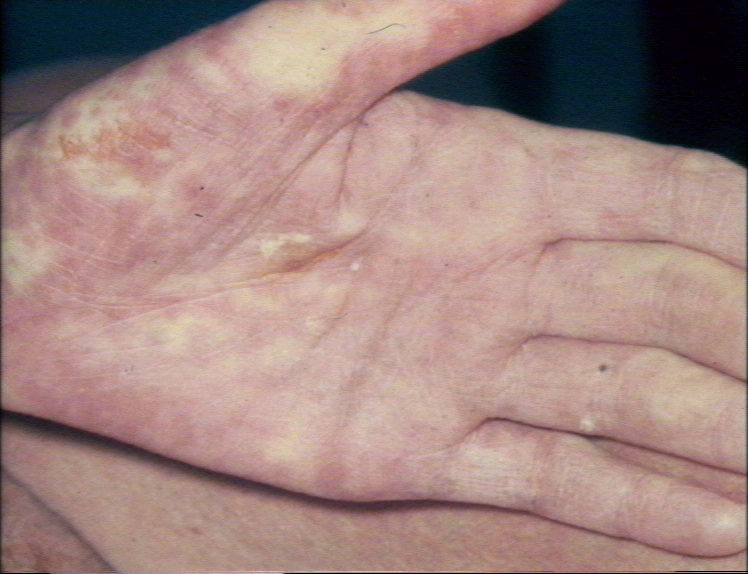 Acute graft-versus-host reaction with vivid palmar erythema ©
Bristol Biomedical Image Archive. Used with permission
Acute graft-versus-host reaction with vivid palmar erythema ©
Bristol Biomedical Image Archive. Used with permission
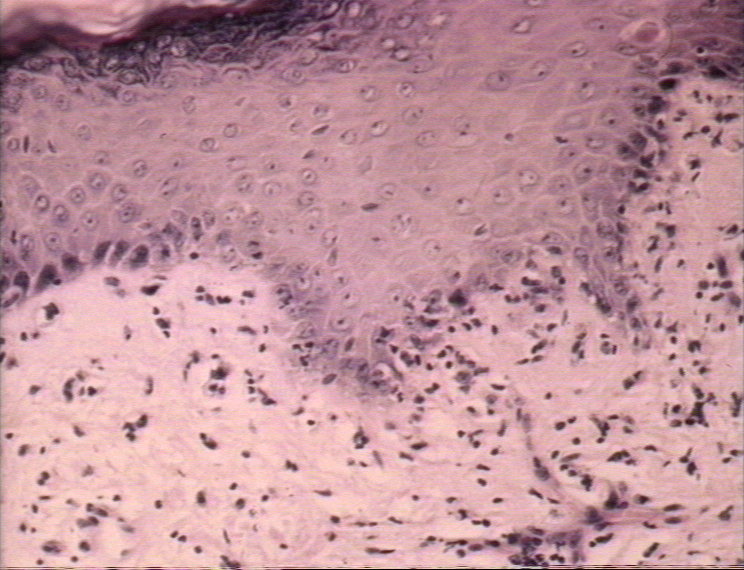
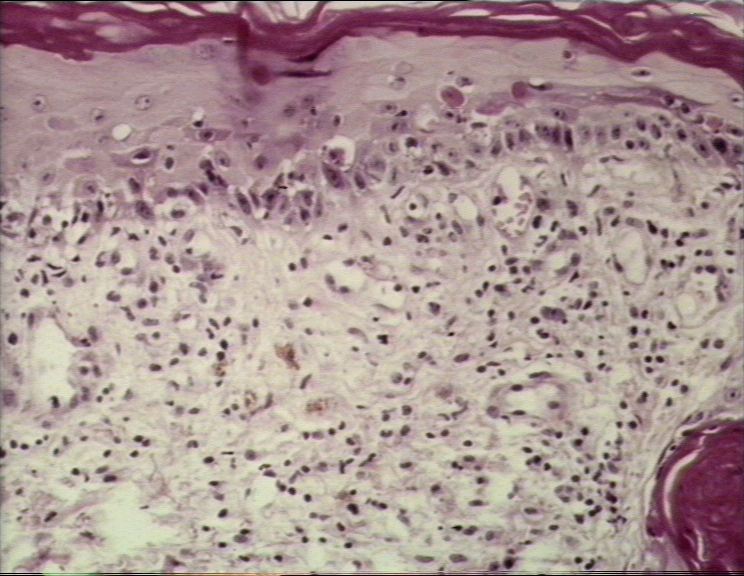 Graft-versus-host reaction; acute basal cell hydropic degeneration with interepidermal necrotic keratinocytes
Graft-versus-host reaction; acute basal cell hydropic degeneration with interepidermal necrotic keratinocytes
© Bristol Biomedical Image Archive. Used with permission
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
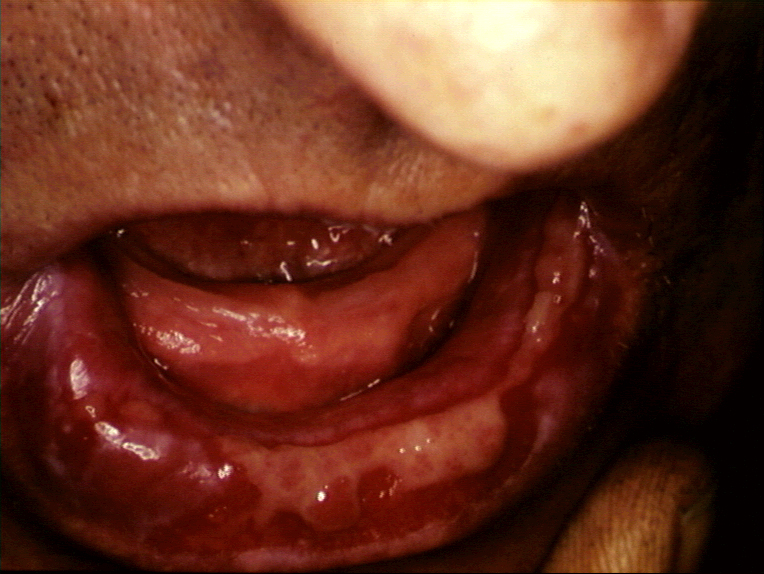
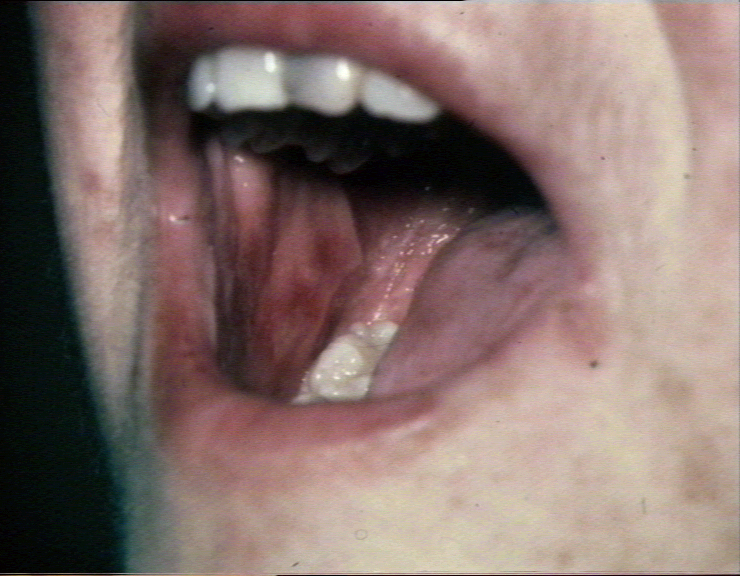 Acute erosions of the buccal mucosa in graft-versus-host reaction
©
Bristol Biomedical Image Archive. Used with permission
Acute erosions of the buccal mucosa in graft-versus-host reaction
©
Bristol Biomedical Image Archive. Used with permission |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
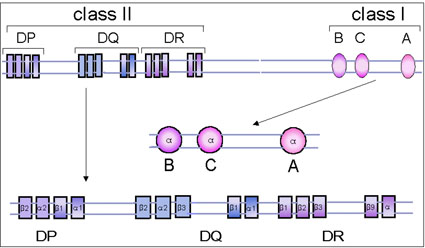 Figure 5 The human MHC gene complex
Figure 5 The human MHC gene complex |
THE MHC GENE COMPLEX The MHC complex contains a number of genes that control several antigens, most of which influence allograft rejection. These antigens (and their genes) can be divided into three major classes: class I, class II and class III. The class I and class II antigens are expressed on cells and tissues whereas as class III antigens are represented on proteins in serum and other body fluids (e.g.C4, C2, factor B, TNF). Antigens of class III gene products have no role in graft rejection.
The human MHC is located on chromosome 6.
The class I gene complex contains three major loci, B, C and A and other undefined minor loci (figure 5). Each major locus codes for a polypeptide; the alpha-chain that contains antigenic determinants, is polymorphic (has many alleles). It associates with beta-2 microglobulin (beta-chain), encoded by a gene outside the MHC complex, and expressed on the cell surface. Without the beta-2 microglobulin, the class I antigen will not be expressed on the cell surface. Individuals with a defective beta-2 microglobulin gene do not express any class I antigen and hence have a deficiency of cytotoxic T cells. Class II MHC The class II gene complex also contains at least three loci, DP, DQ and DR; each of these loci codes for one alpha- and one beta-chain polypeptide which associate together to form the class II antigens. Like the class I antigens, the class II antigens are also polymorphic. The DR locus may contain more than one, possibly four, functional beta-chain genes.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
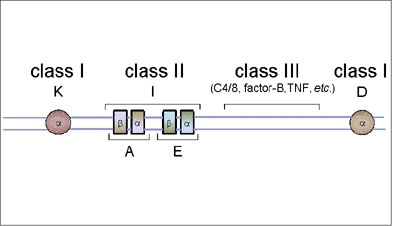 Figure 6A The mouse MHC complex
Figure 6A The mouse MHC complex |
The mouse MHC is located on chromosome 17.
This consists of two major loci, K and D. Unlike the human MHC, the mouse class I gene complexes loci are not together but they are separated by class II and class III genes (Figure 6A). Class II MHC The class II gene complex contains two loci, A and E, each of which code for one alpha and one beta chain polypeptide, which form one class II molecule. The mouse class II gene complex is also known as the I region and the genes in this complex are referred to as Ir (immune response) genes since they determine the magnitude of immune responsiveness of different mouse strains to certain antigens. Products of the A and E loci are also termed IA and IE antigens, collectively known as Ia antigens.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
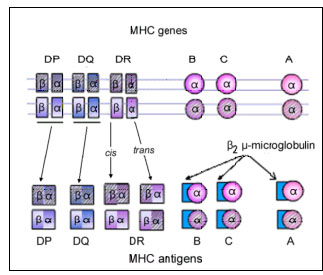 Figure 6B Codominant expression of MHC antigens
Figure 6B Codominant expression of MHC antigens |
MHC ANTIGENS
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
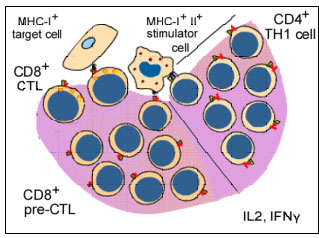 Figure 7 Activation of CTL during MLR
Figure 7 Activation of CTL during MLR |
Another consequence of the MHC antigen and T cell interaction is the induction of cytotoxic T-lymphocytes. When T-lymphocytes are cultured in the presence of allogeneic lymphocytes, in addition to undergoing mitosis (MLR), they also become cytotoxic to cells of the type that stimulated MLR (figure 7). Thus, T-lymphocytes of 'x' haplotype cultured over 5 - 7 days with B lymphocytes of 'y' haplotype will undergo mitosis and the surviving T-lymphocytes become cytotoxic to cells of the 'y' haplotype. The induction of mitosis in MLR requires disparity of only class II antigens whereas the induction of cytotoxic T-lymphocytes (CTL) requires disparity of both class I and class II antigens. However, once cytotoxic cells have been induced, the effector cytotoxic cells recognize only class I antigens to cause cytotoxicity.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
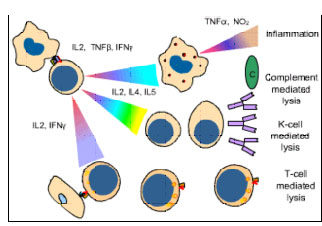 Figure
8 Mechanisms of allograft destruction Figure
8 Mechanisms of allograft destruction |
ALLOGRAFT REJECTION The clinical significance of the MHC is realized in organ transplantation. Cells and tissues are routinely transplanted as a treatment for a number of diseases. However, reaction of the host against allo-antigens of the graft (HVG) results in its rejection and is the major obstacle in organ transplantation. The rejection time of a graft may vary with the antigenic nature of the graft and the immune status of the host and is determined by the immune mechanisms involved (Figure 8 and Table 1).
This occurs in instances when the
recipient has preformed high titer antibodies. A graft may show signs of
rejection within minutes to hours due to immediate reaction of antibodies and
complement. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Transplantation of a second graft, which shares a significant number of antigenic determinants with the first one, results in a rapid (2 - 5 days) rejection. It is due to presence of T-lymphocytes sensitized during the first graft rejection. Accelerated rejection is mediated by immediate production of lymphokines, activation of monocytes and macrophages, and induction of cytotoxic lymphocytes.
The normal reaction that follows the
first grafting of a foreign transplant takes 1 - 3 weeks. This is known as acute
rejection and is mediated by T lymphocytes sensitized to class I and class II
antigens of the allograft, elicitation of lymphokines and activation
of monocytes and macrophages. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
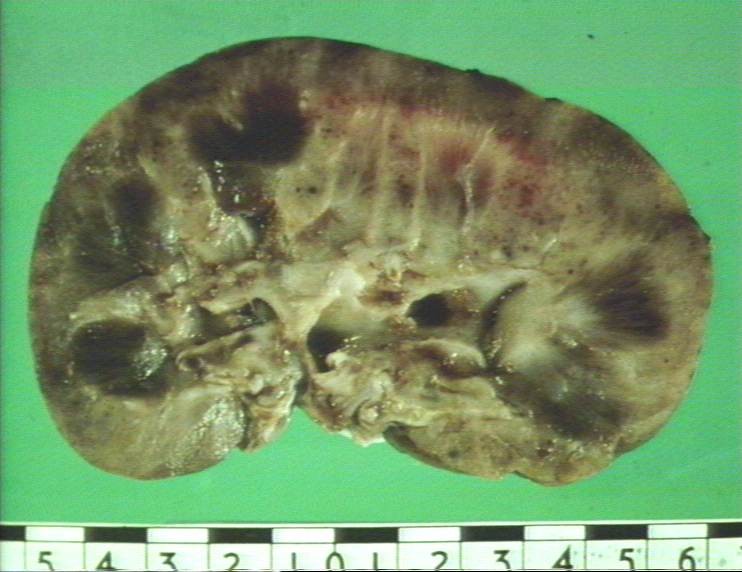 Figure
9A Figure
9A Kidney Transplantation Graft Rejection © Bristol Biomedical Image Archive. Used with permission |
Some grafts may survive for months or even years, but suddenly exhibit symptoms of rejection. This is referred to as chronic rejection, the mechanism of which is not entirely clear. The hypotheses are that this may be due infection, causes which led to failure of the first organ, loss of tolerance induced by the graft, etc. Fetus as an Allograft The fetus in an out-bred mammalian species bears antigens
derived from both the father and the mother. Thus, truly, the fetus is an
allograft and the mother should normally recognize the fetus as foreign and
reject the fetus. Nonetheless, such rejections seldom occur. Thus, mammals
have adapted in a way that allows implantation of their embryos in the
mother's womb and their subsequent survival. There are multiple mechanisms
that play a role, of which the most important being the unique structure and
function of placenta. There are certain locations in the body in which allografts are not readily rejected. These include the brain, anterior chamber of the eye, testis, renal tubule, uterus, etc. This stems from the fact that such sites may lack of good lymphatic drainage. Also, such tissues may express molecules such as Fas ligand that kills any immune cell that may come in contact with these tissues. Additionally, such tissues, may have other immune suppressor mechanisms. Similarly, there are some tissues that can be transplanted without matching and without being rejected. Such tissues are called immunologically privileged tissues. Corneal graft is an excellent example that enjoys the highest success rate of any form of organ transplantation. The low incidence of graft rejection is impressive despite the fact that HLA antigen matching of donor and recipient is not normally performed. There are many explanations as to why such grafts are accepted. The avascularity of the graft bed prevents corneal alloantigens from reaching the regional lymphoid tissues. Also, the corneal antigens may be masked. Together, such mechanisms fail to activate the immune system of the recipient.
PROCEDURES TO ENHANCE GRAFT SURVIVAL In clinical practice, the most successful transplantation programs have been with kidneys and corneas. However, other organs are being transplanted with increasing frequency. The success in these programs has been due to a better understanding of immunological mechanisms, definition of MHC antigens and development of more effective immunosuppressive agents. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Figure
9B Figure
9BKidney Transplantation Chronic Graft Rejection © Bristol Biomedical Image Archive. Used with permission |
Based on extensive experiences with renal transplants, certain guidelines can be followed in donor selection and recipient preparation for most organ transplants. The most important in donor selection is the MHC identity with the recipient; an identical twin is the ideal donor. Grafts from an HLA-matched sibling have 95-100% chance of success. One haplotype-identical parent or sibling must match at the HLA D region. A two haplotype-distinct donor with a reasonable match for D-region antigen can also be used. Organs from a two or one DR matched cadaver have been used also with some success. In every case, an ABO compatibility is essential. Recipient preparation The recipient must be infection-free and must not be hypertensive. One to five transfusions of 100-200 ml whole blood from the donor at 1-2 week intervals improves the graft survival and is practiced when possible. Immunosuppression Immunosuppressive therapy is most
essential part of allo-transplantation. The most recent and effective family of
agents is cyclosporin A, FK-506 (tacrolimus) and rapamycin. Cyclosporin A and
FK506 inhibit IL-2 synthesis following Ag-receptor binding whereas rapamycin
interferes with signal transduction following IL2 - IL2 receptor interaction. Thus, all
three agents block T cell proliferation in response to antigen. Other
chemical agents used to prevent graft rejection and their modes of action have
been listed in Table 2. Whole body irradiation is used in leukemia patients
before bone marrow transplantation. Antisera against T cells (anti-thymocyte
globulin: ATG) or their surface antigens (CD3, CD4, CD45 on activated T-cells,
CD25:IL-2 receptors) are being used also to achieve immunosuppression (Table 2). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
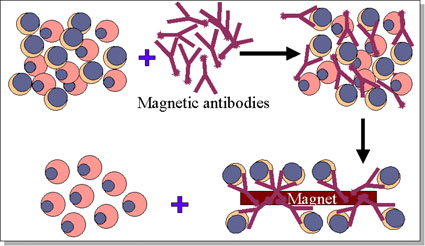 Figure 10 Removal of T cells from marrow graft
Figure 10 Removal of T cells from marrow graft |
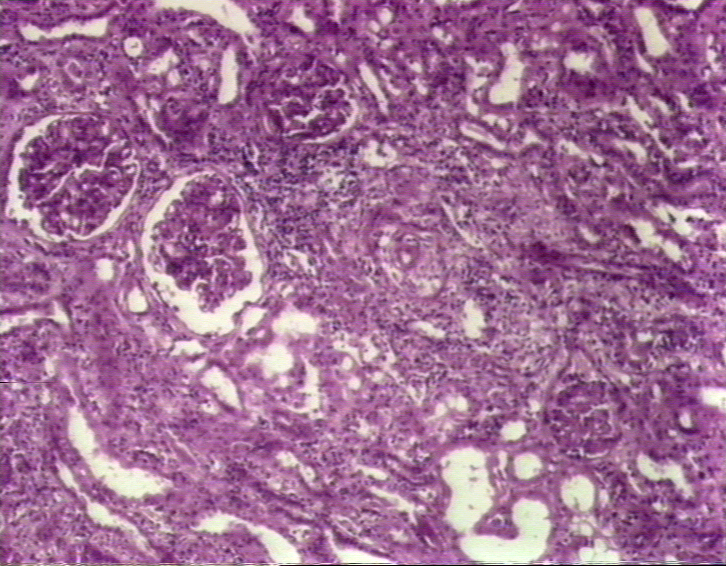
In bone marrow transplantation, the most crucial factor in donor selection is class II MHC compatibility. Once again an identical twin is the ideal donor. From poorly matched grafts, T lymphocytes can be removed using monoclonal antibodies (figure 10). The recipient must be immunosuppressed. Malignant cells must be eliminated from the recipient blood (in case of blood-borne malignancies). Methotrexate, cyclosporin and prednisone are often used to control GVH disease. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Corneal grafts do not contain D region antigens and consequently survival is frequent. Small grafts are better and corticosteroids are helpful. Skin allografts have a very poor success rate and immunosuppressive therapy is relatively ineffective. Nevertheless, they are often used to provide a temporary covering to promote healing in severe skin damage. Indeed, there will be no rejection if the host and donor are perfectly matched (identical twins) or the recipient is tolerant to the donor MHC antigens (bone marrow chimeras).
A number of diseases have been found to occur at a higher frequency in individuals with certain MHC haplotypes. Most prominent among these are ankylosing spondylitis (B27), celiac disease (DR3) and Reiter's syndrome (B27). Other diseases associated with different specificities of the MHC are listed in Table 3. No definite reason is known for this association. However, several hypotheses have been proposed: antigenic similarity between pathogens and MHC, antigenic hypo- and hyper-responsiveness controlled by the class II genes are included among them.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
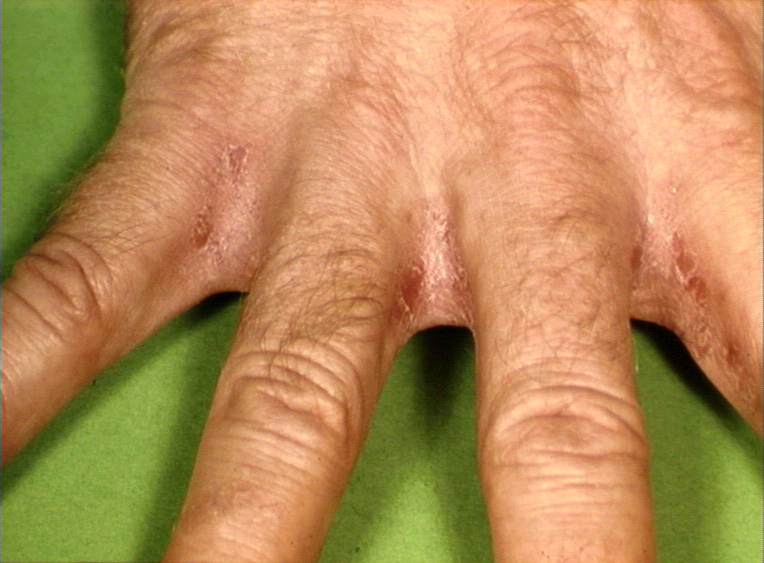 Figure 11 Psoriasis of the hand
© Bristol Biomedical
Image Archive. Used with permission
Figure 11 Psoriasis of the hand
© Bristol Biomedical
Image Archive. Used with permission |
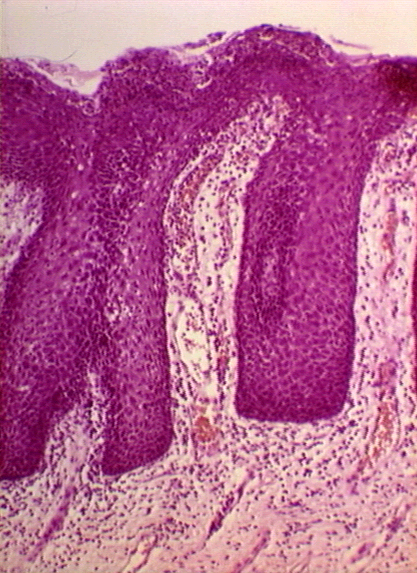 Psoriasis © Bristol Biomedical Image Archive.
Used with permission
Psoriasis © Bristol Biomedical Image Archive.
Used with permission
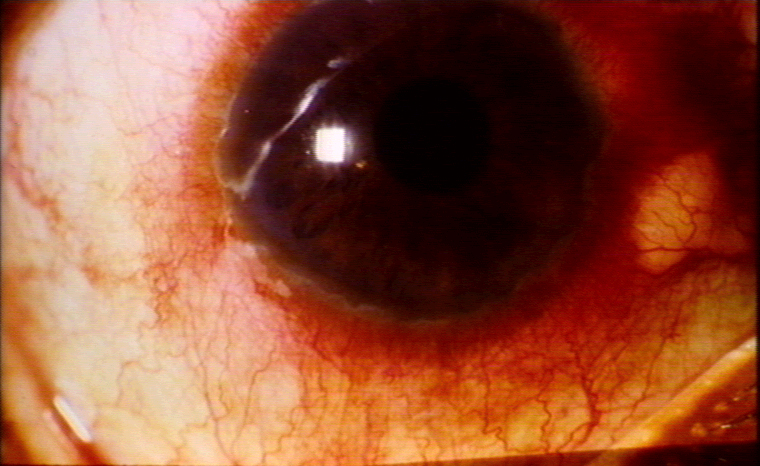 Uveitis © Bristol Biomedical Image Archive. Used
with permission
Uveitis © Bristol Biomedical Image Archive. Used
with permission |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||