|
x |
x |
|
 |
 |
|
INFECTIOUS
DISEASE |
BACTERIOLOGY |
IMMUNOLOGY |
MYCOLOGY |
PARASITOLOGY |
VIROLOGY |
|
VIETNAMESE |
IMMUNOLOGY - CHAPTER EIGHT
ANTIBODY FORMATION
Dr Gene Mayer
Emertius Professor of Pathology, Microbiology and Immunology
University of South Carolina
|
|
TURKISH |
|
FRANCAIS |
|
PORTUGUES |
|
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|
|

 |
|
Logo image
© Jeffrey Nelson, Rush University, Chicago, Illinois and
The MicrobeLibrary |
|
|
|
|
TEACHING OBJECTIVES
To describe general
characteristics of the specific immune response
To compare and contrast
primary and secondary antibody responses
To describe the molecular
events involved in class switching and membrane immunoglobulin expression |
GENERAL
CHARACTERISTICS OF THE ANTIBODY RESPONSE
Self / non-self
discrimination
One characteristic feature of the specific immune
system is that it normally distinguishes between self and non-self
and only reacts against non-self.
Memory
A second
feature of the specific immune response is that it demonstrates memory.
The immune system "remembers" if it has seen an antigen before
and it reacts to secondary exposures to an antigen in a manner different
than after a primary exposure. Generally only an exposure to the same
antigen will illicit this memory response.
Specificity
A
third characteristic feature of the specific immune system is that there
is a high degree of specificity in its reactions. A response to a
particular antigen is specific for that antigen or a few closely related
antigens.
N.B. These are characteristic
of all specific immune responses.
|
|
KEY WORDS
Equilibrium
phase
Primary
response
Steady state
phase
Class
switching
Catabolic decay
phase
Lag/inductive
phase
Decline
phase
Immune elimination
phase
Log
phase
Secondary/anamnestic
response
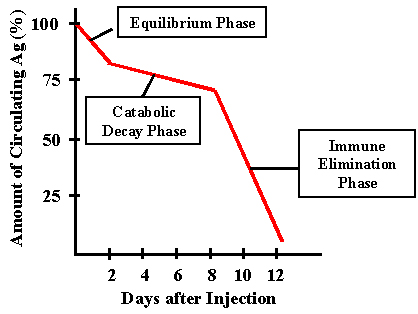
 Figure 1
Figure 1 |
ANTIBODY
FORMATION
Fate
of the immunogen
Clearance after primary
injection
The kinetics of antigen clearance from the body after a primary
administration is depicted in Figure 1.
Equilibrium phase
The
first phase is called the equilibrium or equilibration phase. During this
time the antigen equilibrates between the vascular and extravascular
compartments by diffusion. This is normally a rapid process. Since
particulate antigens don't diffuse, they do not show this phase.
Catabolic decay phase
In
this phase the host's cells and enzymes metabolize the antigen. Most of
the antigen is taken up by macrophages and other phagocytic cells. The
duration will depend upon the immunogen and the host.
Immune elimination phase
In this phase, newly synthesized antibody combines with the antigen
producing antigen/antibody complexes which are phagocytosed and degraded.
Antibody appears in the serum only after the immune elimination phase is
over.
Clearance after secondary
injection
If there is circulating antibody in the serum, injection of
the antigen for a second time results in a rapid immune elimination. If the
is no circulating antibody then injection of the antigen for a second time
results in all three phases but the onset of the immune elimination phase is
accelerated.
|
 Figure
2
Figure
2 |
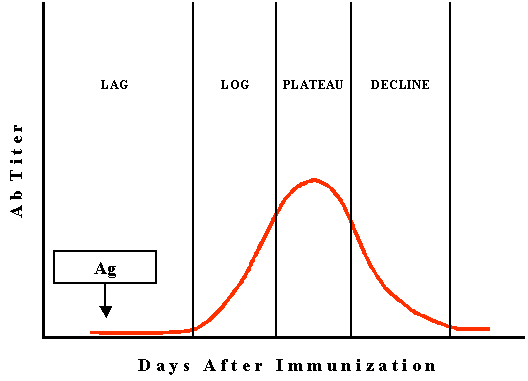
Kinetics
of antibody responses to T-dependent Antigen
Primary (1o)
Antibody response
The kinetics of a primary antibody response to an antigen
is illustrated in Figure 2.
Inductive, latent or lag
phase
In this phase, the antigen is recognized as foreign and the cells begin
to proliferate and differentiate in response to the antigen. The duration
of this phase will vary depending on the antigen but it is usually 5 to 7
days.
Log or Exponential Phase
In this phase, the antibody concentration increases exponentially as the B cells
that were stimulated by the antigen differentiate into plasma cells which
secrete antibody.
Plateau or steady-state phase
In this phase, antibody synthesis is balanced by antibody decay so that there
in no net increase in antibody concentration.
Decline or decay phase
In this phase, the rate of antibody degradation exceeds that of antibody synthesis and
the level of antibody falls. Eventually the level of antibody may reach base line
levels.
|
 Figure
3
Figure
3 |
Secondary (2o),
memory or anamnestic response (Figure 3)
Lag phase
In a secondary
response, there is a lag phase by it is normally shorter than that observed
in a primary response.
Log phase
The log phase
in a secondary response is more rapid and higher antibody levels are achieved.
Steady state phase
Decline phase
The
decline phase is not as rapid and antibody may persist for months, years or even
a lifetime.
Specificity of primary
and secondary responses
Antibody elicited in response to an
antigen is specific for that antigen, although it may also cross react with
other antigens which are structurally similar to the eliciting antigen. In
general secondary responses are only elicited by the same antigen
used in the primary response. However, in some instances a closely related
antigen may produce a secondary response, but this is a rare exception.
|
 Figure
4
Figure
4
|
Qualitative changes in
antibody during primary and secondary responses
Immunoglobulin class variation
In the primary response, the major class of antibody produced is IgM whereas in the
secondary response it is IgG (or IgA or IgE) (Figure 4). The antibodies that
persist in the secondary response are the IgG antibodies.
|
 Figure 5
Figure 5 |
Affinity
The
affinity of the IgG antibody produced increases progressively during the response,
particularly after low doses of antigen (Figure 5). This is referred to as
affinity maturation. Affinity maturation is most pronounced after secondary
challenge with antigen.
|
 Figure 6
Figure 6 |
One explanation for affinity
maturation is clonal selection as illustrated in Figure 6. A second
explanation for affinity maturation is that, after a class switch has
occurred in the immune response, somatic mutations occur which fine tune the
antibodies to be of higher affinity. There is experimental evidence for this
mechanism, although it is not known how the somatic mutation mechanism is
activated after exposure to antigen.
Avidity
As a
consequence of increased affinity, the avidity of the antibodies increases
during the response.
Cross-reactivity
As
a result of the higher affinity later in the response, there is also an
increase in detectible cross reactivity. An explanation for why increasing
affinity results in an increase in detectible cross reactivity is
illustrated by the following example.
|
| |
|
Affinity of Ab for Ag |
| |
Early |
Late |
| Immunizing Ag |
10-6
|
10-9
|
|
+ |
++ |
| Cross reacting Ag |
10-3
|
10-6
|
|
- |
+ |
| |
If a minimum affinity of 10-6
is needed to detect a reaction, early in an immune response the reaction of a
cross reacting antigen with an affinity of 10-3 will not be
detected. However, late in a response when the affinities increase 1000 fold,
the reaction with both the immunizing and cross reacting antigens will be
detected.
|
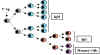 Figure
7
Figure
7 |
Cellular events during
primary
and secondary responses to T-dependent antigen
Primary response
(Figure 7)
Lag phase
Clones of T and
B cells with the appropriate antigen receptors bind antigen, become
activated and begin to proliferate. The expanded clones of B cells
differentiate into plasma cells which begin to secrete antibody.
Log phase
The plasma
cells initially secrete IgM antibody since the Cμ heavy
chain gene is closest to the rearranged VDJ gene. Eventually some B cells
switch from making IgM to IgG, IgA or IgE. As more B cells proliferate and
differentiate into antibody secreting cells the antibody concentration
increases exponentially.
Stationary phase
As
antigen is depleted, T and B cells are no longer activated. In addition,
mechanisms which down regulate the immune response come into play.
Furthermore, plasma cells begin to die. When the rate of antibody
synthesis equals the rate of antibody decay the stationary phase is
reached.
Decline phase
When no
new antibody is produced because the antigen is no longer present to
activate T and B cells and the residual antibody slowly is degraded, the
decay phase is reached.
|
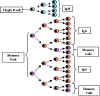 Figure 8
Figure 8
 Figure
9
Figure
9 |
Secondary response
(Figure 8)
Not all of the T and B cells
that are stimulated by antigen during primary challenge with antigen die.
Some of them are long lived cells and constitute what is refer to as the
memory cell pool. Both memory T cells and memory B cells are produced and
memory T cells survive longer than memory B cells. Upon secondary
challenge with antigen not only are virgin T and B cells activated, the
memory cells are also activated and thus there is a shorter lag time in
the secondary response. Since there is an expanded clone of cells being
stimulated the rate of antibody production is also increased during the
log phase of antibody production and higher levels are achieved. Also,
since many if not all of the memory B cells will have switched to IgG (IgA
or IgE) production, IgG is produced earlier in a secondary response.
Furthermore since there is an expanded clone of memory T cells which can
help B cells to switch to IgG (IgA or IgE) production, the predominant
class of Ig produced after secondary challenge is IgG (IgA or IgE).
Ab response to
T-independent antigen
Responses to T-independent antigen are characterized by
the production of almost exclusively IgM antibody and no secondary response.
Secondary exposure to the antigen results in another primary response to the
antigen as illustrated in Figure 9.
|
 Figure
10
Figure
10 |
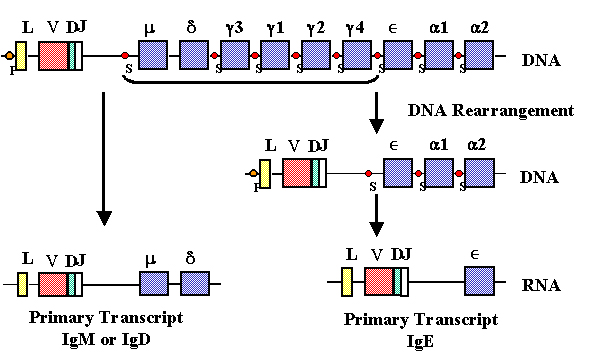
Class
switching
During an antibody response
to a T-dependent antigen a switch occurs in the class of Ig produced from
IgM to some other class (except IgD). Our understanding of the structure
of the immunoglobulin genes, helps explain how class switching occurs
(Figure 10).
During class switching
another DNA rearrangement occurs between a switch site (Sμ)
in the intron between the rearranged VDJ regions and the Cμ
gene and another switch site before one of the other heavy chain constant
region genes. The result of this recombination event is to bring the VDJ
region close to one of the other constant region genes, thereby allowing
expression of a new class of heavy chain. Since the same VDJ gene is
brought near to a different C gene and since the antibody specificity is
determined by the hypervariable regions within the V region, the antibody
produced after the switch occurs will have the same specificity as before.
Cytokines secreted by T
helper cells can cause the switch to certain isotypes.
|
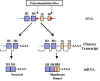 Figure 11
Figure 11 |
Membrane and secreted
immunoglobulin
The specificity of membrane
immunoglobulin on a B cell and the Ig secreted by the plasma cell progeny
of a B cell is the same. An understanding of how the specificity of
membrane and secreted Ig from an individual B cell can be the same comes
from an understanding of immunoglobulin genes (Figure 11).
There are two potential polyA
sites in the immunoglobulin gene. One after the exon for the last heavy
chain domain and the other after the exons that code for the trans-
membrane domains. If the first polyA site is used, the pre-mRNA is
processed to produce a secreted protein. If the second polyA site is used,
the pre-mRNA is processed to produce a membrane form of the immunoglobulin.
However, in all cases the same VDJ region is used and thus the specificity
of the antibody remains the same. All C regions genes have these
additional membrane pieces associated with them and thus after class
switching other classes of immunoglobulins can be secreted or expressed on
the surface of B cells.
|
|
|
 Return to the Immunology Section of Microbiology and Immunology On-line Return to the Immunology Section of Microbiology and Immunology On-line
This page last changed on
Wednesday, August 30, 2017
Page maintained by
Richard Hunt
|
 Figure
2
Figure
2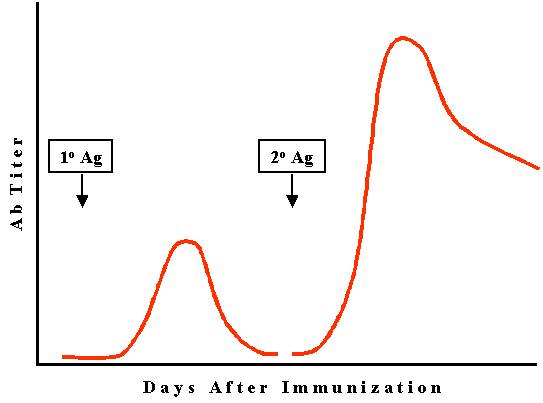 Figure
3
Figure
3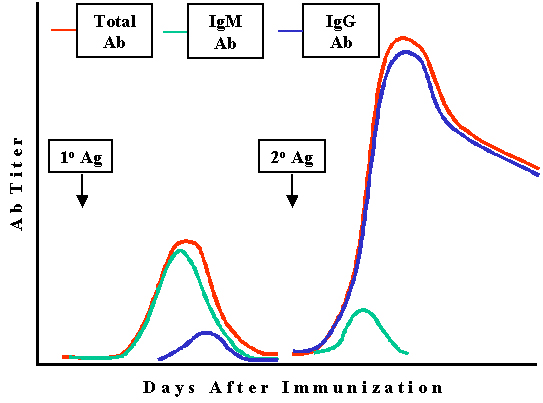 Figure
4
Figure
4
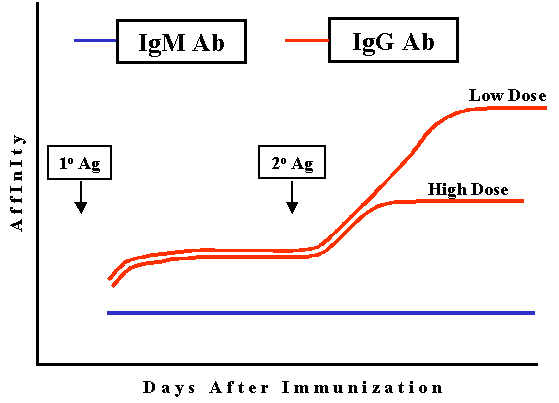 Figure 5
Figure 5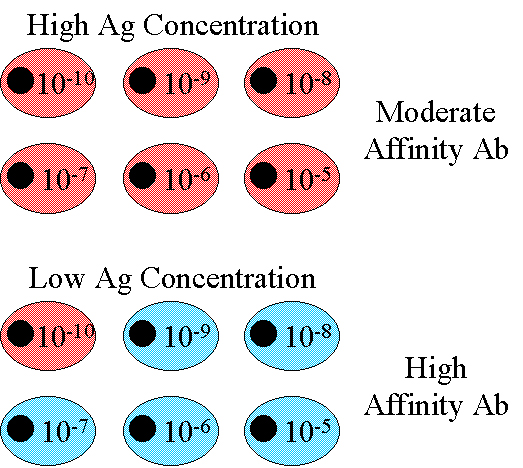 Figure 6
Figure 6 Figure
7
Figure
7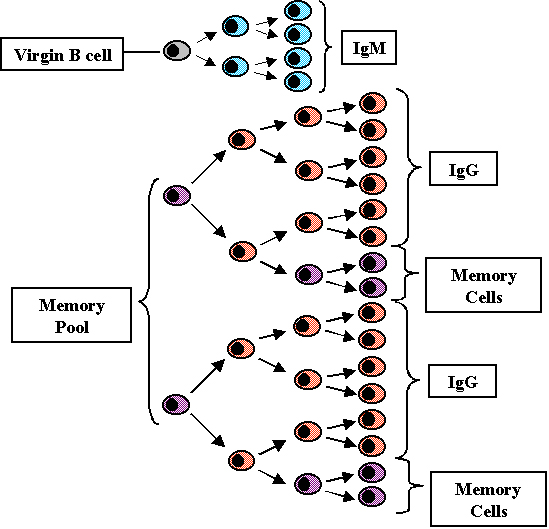 Figure 8
Figure 8
 Figure
10
Figure
10 Figure 11
Figure 11