|
|
|
|
 |
 |
|
DOENÇAS
INFECCIOSAS |
BACTERIOLOGIA |
IMUNOLOGIA |
MICOLOGIA |
PARASITOLOGIA |
VIROLOGIA |
|
|
IMUNOLOGIA - CAPÍTULO OITO
FORMAÇÃO
DE ANTICORPOS
Dr.
Gene Mayer
Professor Emeritus
University of South Carolina School of Medine
Tradução:
PhD. Myres Hopkins
|
|
EM INGLÊS
|
|
|
|
Dê a
sua opinião
CONTATO |
|
BUSCA |
|
E-MAIL
DR MYRES HOPKINS |
|
ESCOLA DE MEDICINA DA
UNIVERSIDADE DA CAROLINA DO SUL |
|

 |
|
|
|
OBJETIVOS
Descrever as características gerais da resposta imune específica
Comparar e diferenciar as respostas imunes primárias e secundárias
Descrever os eventos moleculares envolvidos na mudança de classe e na
expressão de imunoglobulina de membrana |
CARACTERÍSTICAS
GERAIS DA RESPOSTA POR ANTICORPO
Discriminação entre o próprio/não próprio
Um aspecto característico do sistema imune específico é que ele
normalmente distingue entre o próprio e o não próprio e reage apenas
contra o não próprio.
Memória
Um segundo aspecto da resposta imune específica é que ele demonstra
memória. O sistema imune “se lembra” de que já viu um antígeno antes e
reage a uma segunda exposição do antígeno de maneira diferente da
primeira exposição. Geralmente apenas uma exposição ao mesmo antígeno
irá iniciar a resposta por memória.
Especificidade
Um terceiro aspecto característico do sistema imune específico é que há
um elevado grau de especificidade em suas reações. A resposta a um
antígeno particular é específica para aquele antígeno ou para alguns
antígenos estreitamente relacionados.
N.B. Estas são características de todas as respostas
imunes específicas.
|
|
PALAVRAS CHAVE
Fase de equilíbrio
Resposta primária
Fase estacionária
Mudança de classe
Fase de decaimento catabólico
Fase latente/indutiva
Fase de declínio
Fase de eliminação imune
Fase log
Resposta secundária /anaméstica
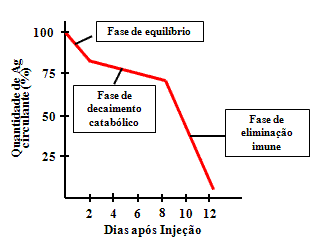
 Figura 1
Figura 1 |
FORMAÇÃO DO
ANTICORPO
Destino do imunógeno
Eliminação após injeção primária
A cinética de eliminação do antígeno do corpo após a primeira
administração é mostrada na Figura 1.
-
Fase de equilíbrio
A primeira fase é chamada de fase de
equíbrio ou equilibração. Durante este tempo o antígeno se
equilibra entre os compartimentos vasculares e extravasculares
por difusão. Este é normalmente um processo rápido. Uma vez que
antígenos particulados não difundem, eles não têm esta fase.
-
Fase de decaimento catabólico
Nesta fase as células
hospedeiras e enzimas metabolizam o antígeno. A maioria dos
antígenos é englobada por macrófagos e outras células
fagocíticas. A duração vai depender do imunógeno e do
hospedeiro.
-
Fase de eliminação imune
Nesta fase os anticorpos recém
sintetizados se combinam com o antígeno produzindo complexos
antígeno/anticorpo que são fagocitados e degradados. Os
anticorpos aparecem no soro somente após concluida a fase de
eliminação imune.
Eliminação após injeção secundária
Se há anticorpo circulando no soro a injeção de um antígeno pela
segunda vez resulta em uma eliminação imune rápida. Se não há
anticorpo circulando então a injeção do antígeno pela segunda vez
resulta em todas as três fases, mas o ataque da fase de eliminação é
acelerado.
|
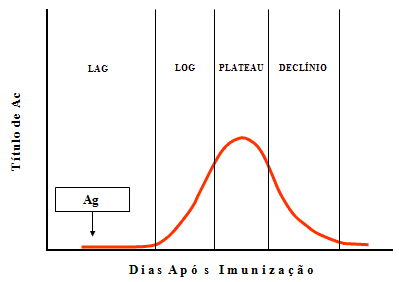
 Figura
2
Figura
2 |
Cinética de respostas de anticorpos a um antígeno T-dependente
Resposta primária (1ria) de anticorpos
A cinética de uma resposta primária de anticorpos a um antígeno é
ilustrada na Figura 2.
-
Fase indutiva e latente ou lag – Nesta fase o antígeno é reconhecido
como estranho e as células começam a proliferar e a se diferenciar
em resposta ao antígeno. A duração desta fase irá variar dependendo
do antígeno mas é normalmente de 5-7 dias.
-
Fase log ou Exponencial – Nesta fase a concentração do anticorpo
aumenta exponencialmente pois as células B que foram estimuladas
pelo antígeno se diferenciam em plasmócitos que secretam anticorpos.
-
Fase de plateau ou de equiliíbrio estacionário – Nesta fase a
síntese de anticorpos é equilibrada com o decaimento de anticorpos,
de forma que não há incremento na concentração de anticorpos.
-
Fase de declínio ou de decaimento – Nesta fase a
taxa de degradação de anticorpos excede a taxa de síntese e o nível
de anticorpos cai. Eventualmente o nível atinge os níveis
constitutivos.
|
 Figura
3
Figura
3 |
Resposta secundária (2ria), de
memória ou anamnésica
(Figura 3)
-
Fase lag
Em uma resposta secundária há
uma fase lag mas é normalmente mais curta do que a observada
na resposta primária.
-
Fase log – A fase log em uma resposta
secundária é mais rápida e atinge níveis mais elevados de
anticorpos.
-
Fase de equilibrio estacionário
-
Fase de declínio – A fase de declínio não
é tão rápida e o anticorpo pode persistir durante meses,
anos ou mesmo durante a vida toda.
Especificidade das respostas 1ria e 2ria
O anticorpo produzido em resposta a um antígeno é
específico para aquele antígeno, embora possa também fazer reação
cruzada com outros antígenos que sejam estruturalmente semelhantes
ao antígeno que o originou. Em geral respostas secundárias são
emitidas pelo mesmo antígeno usado na resposta primária. Entretanto,
em algumas situações um antígeno estreitamente relacionado pode
produzir uma resposta secundária, mas esta é uma rara exceção.
|
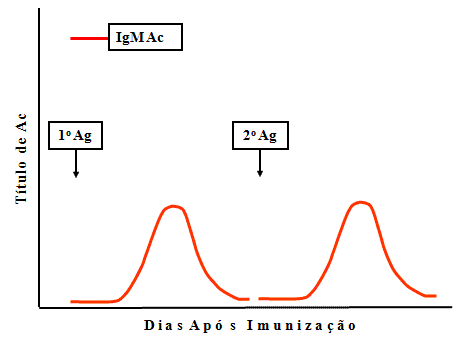
 Figura
4
Figura
4
|
Mudanças qualitativas no anticorpo durante as respostas 1ria
e 2ria
Variação de classe de Ig
Na resposta primária a classe principal de
anticorpo produzido é a IgM, enquanto que na resposta secundária
é IgG (ou IgA ou IgE) (Figura 4). Os anticorpos que persistem na
resposta secundária são os anticorpos IgG.
|
 Figura 5
Figura 5 |
Afinidade
A afinidade de Ac IgG produzidos aumenta
progressivamente durante a resposta, particularmente após doses
pequenas do antígeno (Figura 5). Isso é chamado de maturação por
afinidade. Maturação por afinidade é mais pronunciada após uma
segunda provocação pelo antígeno.
|
 Figura 6
Figura 6 |
Uma
explicação para a maturação por afinidade é a seleção clonal ilustrada
na Figura 6. Uma segunda explicação para a maturação por afinidade é que
após ocorrida a mudança de classe na resposta imune, mutações somáticas
ocorrem, o que aprimora os anticorpos para se tornarem de afinidade mais
elevada. Há evidência experimental para este mecanismo, ambora não se
saiba como o mecanismo de mutação somática é ativado após exposição ao
antígeno.
Avidez
Como consequência do aumento de afinidade, a avidez dos
anticorpos aumenta durante a resposta.
Reatividade-cruzada
Como resultado da afinidade tardia aumentada na
resposta, também ocorre um aumento na reatividade cruzada detectável.
Uma explicação para o porque do aumento da afinidade levando a um
aumento da reatividade cruzada detectável está ilustrado no seguinte
exemplo.
|
| |
|
Afinidade do Ac pelo Ag |
| |
Inicial |
Tardia |
|
Ag Imunizante |
10-6
|
10-9
|
|
+ |
++ |
|
Ag de reação cruzada |
10-3
|
10-6
|
|
- |
+ |
| |
Se um mínimo de afinidade de 10-6 é
necessária para detectar a reação, inicialmente em uma resposta
imune a reação de um antígeno de reação cruzada com uma
afinidade de 10-3 não será detectada. Entretanto,
mais tarde na resposta, quando a afinidade aumentar 1000 vezes,
a reação com ambos os antígenos imunizante e de reação cruzada
será detectada.
|
 Figura
7
Figura
7 |
Eventos celulares durante as respostas 1rias e 2rias
a antígenos T-independentes
Resposta primária
(Figura 7)
-
Fase lag
Clones de células T e B com os receptores apropriados de
antígenos, tornam´se ativados e começam a proliferar. Os clones
expandidos de células B se diferenciam em plasmócitos que começam a
secretar anticorpos.
-
Fase log
Os plasmócitos inicialmente secretam anticorpo IgM uma
vez que o gene de cadeia pesada Cμ está mais perto do
gene VDJ rearranjado. Eventualmente algumas células B trocam a
produção de IgM pela produção de IgG, IgA ou IgE. À medida que mais
células B proliferam e se diferenciam em células secretoras de
anticorpos a concentração de anticorpos aumenta exponencialmente.
-
Fase estacionária
À medida que o antígeno é esgotado, as células T
e B não são mais ativadas. Em adição a isso, mecanismos que regulam
negativamente a resposta imune começam a participar. Além do mais,
plasmócitos começam a morrer. Quando a taxa de síntese de anticorpo
se iguala à taxa de decaimento, a fase estacionária é atingida.
-
Fase de declínio
Quando nenhum anticorpo novo é
produzido porque o antígeno não está mais presente para ativar
células T e B e o anticorpo residual é paulatinamente degradado, a
fase de decaimento é atingida.
|
 Figura 8
Figura 8
 Figura
9
Figura
9 |
Resposta secundária
(Figura 8)
Nem
todas as células T e B que são estimuladas pelo antígeno durante a
primeira ameaça do antígeno morrem. Algumas delas são células de vida
longa e constituem o que referimos como pool de células de memória.
Tanto células de memória T como células de memória B são produzidas e
células de memónia T sobrevivem mais tempo do que células de memória B.
Numa ameaça secundária pelo antígeno não somente células virgens T e B
são ativadas, como também células de memória e porisso há um tempo de
latência menor na resposta secundária. Uma vez que há um clone expandido
de células sendo estimuladas a taxa de produção de anticorpos também é
aumentada durante a fase log da produção de anticorpos e níveis mais
elevados são atingidos. Igualmente, visto que muitas senão todas as
células de memória B terão mudado para a produção de IgG (IgA ou IgE),
IgG é produzido mais cedo na resposta secundária. Além disso, uma vez
que há um clone expandido de células de memória T que podem ajudar as
células B a mutadarem para a produção de IgG (IgA ou IgE), a classe
predominante de Ig produzida após a segunda ameaça é IgG (IgA ou IgE).
Resposta de Ac a antígenos T-independentes
Respostas a antígenos T-independentes são caracterizadas pela produção de
quase que exclusivamente anticorpos IgM e pela ausência de resposta
secundária. Uma exposição secundária ao antígeno resulta em outra resposta
primária ao antígeno como ilustrado na Figura 9.
|
 Figura
10
Figura
10 |
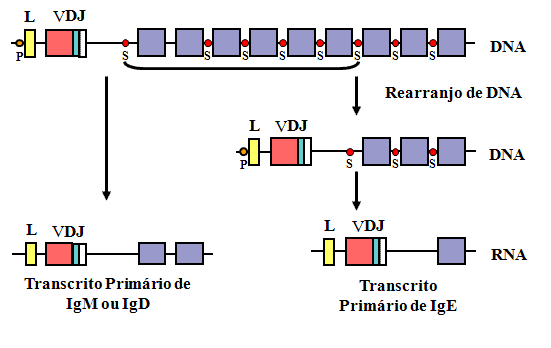
Mudança de classe
Durante uma resposta por anticorpos a um antígeno T-dependente
ocorre uma mudança na classe de Ig produzida de IgM para algumas
outras classes (exceto IgD). Nossa compreensão da estrutura dos
genes de imunoglobulina ajuda a explicar como ocorre a mudança de
classe (Figura 10).
Durante a mudança de classe ocorre outro rearranjo de DNA entre um
sítio de mudança (Sμ) no íntron entre as regiões VDJ
rearranjadas e o gene Cμ e outro sítio de mudança antes
de um dos outros genes de regiões constantes, permitindo dessa forma
a expressão de uma nova classe de cadeia pesada. Uma vez que o mesmo
gene VDJ é colocado junto de um gene C diferente e uma vez que a
especificidade do anticorpo é determinada pelas regiões
hipervariáveis dentro da região V, o anticorpo produzido após a
ocorrência da mudança, terá a mesma especificidade de antes.
Citocinas secretadas pelas células T auxiliares podem
causar a mudança para certos isotipos.
|
 Figura 11
Figura 11 |
Imunoglobulinas de membrana e secretoras
A
especificidade de imunoglobulina de membrana em uma célula B e a Ig
secretada pela progênie de plasmócitos de uma célula B é a mesma.
Uma compreensão de como a especificidade de Ig de membrana e de Ig
secretadas por uma célula B individual podem ser a mesma parte de
uma compreensão dos genes de imunoglobulina (Figura 11).
Há dois sítios poli-A potenciais no gene da
imunoglobulina. Um após o exon para o último domínio de cadeia
pesada e o outro após os exons que codificam para os domínios
trans-membrana. Se o primeiro sítio poli-A é usado, o pré-RNAm é
processado para produzir uma proteína secretora. Se o segundo sítio
poli-A é usado, o pré-RNAm é processado para produzir uma forma
membranar de imunoglobulina. Entretanto, em todos os casos a mesma
região VDJ é usada e porisso a especificidade do anticorpo permanece
a mesma. Todos os genes de região C têm essas partes adicionais de
membrana associadas a eles e assim, após a mudança de classe outras
classes de imunoglobulinas podem ser secretadas ou expressas na
superfície das células B.
|
|
|
 Voltar
à Seção Imunologia do Microbiologia e Imunologia Online
Voltar
à Seção Imunologia do Microbiologia e Imunologia Online
Esta página foi modificada em
Página mantida por
Richard Hunt
Por favor, relate quaisquer problemas para
Richard Hunt
|
 Figura
2
Figura
2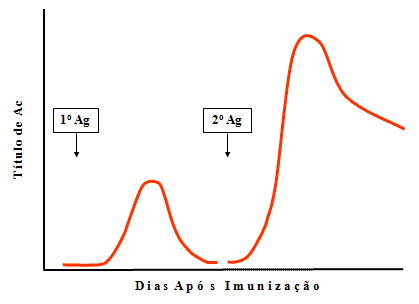 Figura
3
Figura
3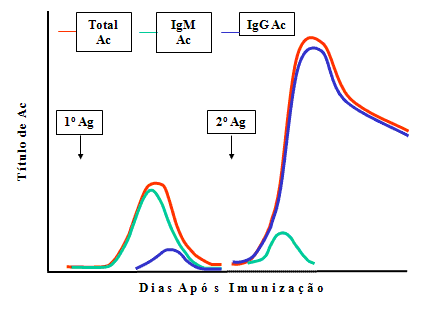 Figura
4
Figura
4
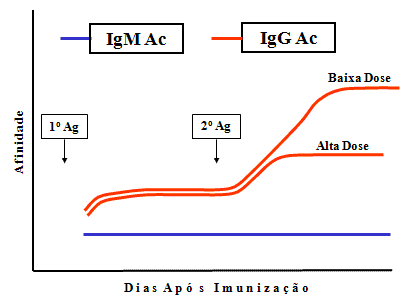 Figura 5
Figura 5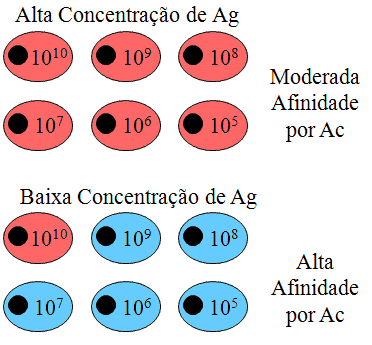 Figura 6
Figura 6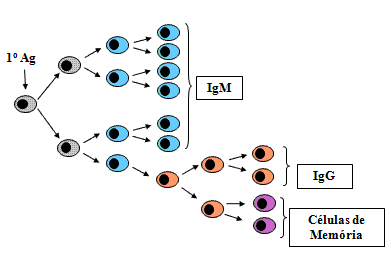 Figura
7
Figura
7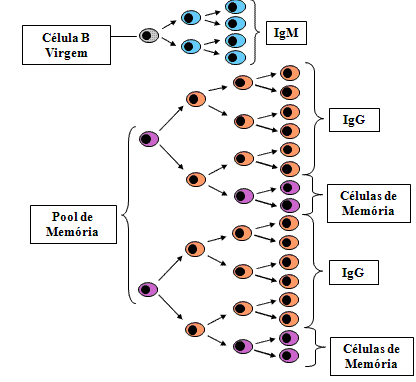 Figura 8
Figura 8
 Figura
10
Figura
10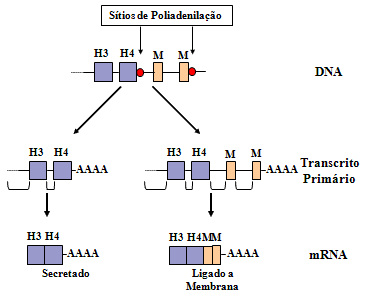 Figura 11
Figura 11