|
|
 |
|||||||||||||||||||||||||||||||
|
|
BACTERIOLOGÍA | INMUNOLOGÍA | MICOLOGÍA | PARASITOLOGÍA | VIROLOGÍA | |||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
|
E-MAIL |
||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
|
PALABRAS CLAVE GlicolisisFermentación Respiración Anaerobia Respiración Aeróbica Anaerobios Facultativos Aerobios obligatorios Anaerobios obligatorios Anaerobios Aerotolerantes Aerobios microaerofilicos Sideróforos Mesófilos Termófilos Psicrofílicos Tiempo de Generación Ciclo de los ácidos Tricarboxilicos (TCA) ó Ciclo de Krebs Ruta del Glioxilato Ubiquinona
|
Las requerimientos para el crecimiento bacteriano incluyen fuentes de energía, carbohidratos (por ejemplo; azúcares y ácidos grasos) e iones metálicos (por ejemplo; fierro). La temperatura óptima, el pH y los requerimientos de la presencia (ó ausencia) de oxígeno son importantes. Requerimientos de Oxígeno Los aerobios obligados deben ser capaces de crecer en presencia de oxígeno y no llevan a cabo fermentación. Los anaerobios obligados no llevan a cabo fosforilación oxidativa. Mas aún, ellos mueren en presencia de oxígeno ya que carecen de ciertas enzimas como la catalasa [la cual rompe el enlace del peróxido de hidrógeno, H2O2, a agua y oxígeno], la peroxidasa [por la cual NADH+ H2O2 se convierten a NAD y O2] y la superóxido dismutasa [por la cual el superóxido, O2; es convertido a H2O2]. Estas enzimas de-toxifican los radicales libres producidos a partir del peróxido de hidrógeno y del oxígeno producidos durante el metabolismo aerobio (en presencia de oxígeno). Los anaeróbios aerotolerantes son bacterias que respiran anaeróbicamentre, pero pueden sobrevivir en presencia de oxígeno. Los anaerobios facultativos pueden llevar cabo tanto la fermentación como la respiración aeróbica. En presencia de oxígeno, la respiración anaeróbica de estos organismos generalmente se apaga y entonces respiran aeróbicamente. Las bacterias micro-aerofílicas crecen bien a bajas concentraciones de oxígeno, pero no resisten altas concentraciones. Requerimientos Nutricionales Estos incluyen fuentes de carbono orgánico, nitrógeno, fósforo, azufre e iones metálicos incluyendo el hierro. Las bacterias secretan moléculas pequeñas que unen el hierro (sideróforos, por ejemplo; enterobactina, micobactina). Los sideróforos (con el hierro unido) son entonces internalizados vía receptores de mebrana por la célula bacteriana. El huésped humano también tiene proteínas transportadoras de hierro (por ejemplo: la transferrina). Por lo tanto las bacterias que compiten con el huésped por el hierro de forma ineficiente no son patógenos exitosos. Temperatura Las bacterias pueden crecer en una variedad temperaturas, desde aquellas cercanas al punto de congelación hasta el punto de ebullición del agua. Las bacterias que crecen mejor a la mitad de este rango se conocen como mesófilos, las cuales incluyen todos los patógenos y oportunistas de humano. Las que viven a temperaturas óptimas altas se conocen como termófilas y las que lo hacen a bajas temperaturas son los psicrófilicos. pH Algunas bacterias crecen mejor a pH neutro, sin embargo ciertas bacterias pueden sobrevivir y crecer de forma constante en condiciones ácidas o alcalinas.
|
|||||||||||||||||||||||||||||||
|
PELICULA
Bacterias y Fisión Binaria
|
||||||||||||||||||||||||||||||||
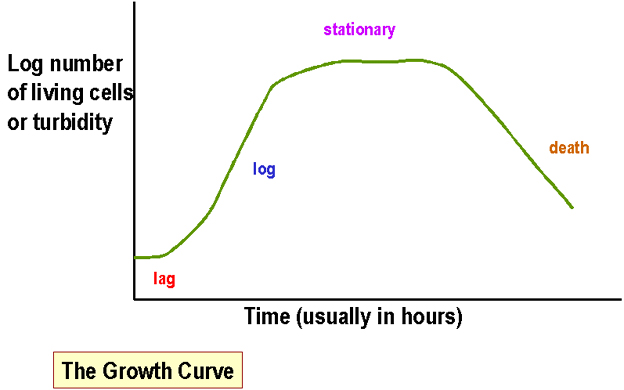 Figura 1. Curva de crecimiento
Figura 1. Curva de crecimiento |
Mediciones de masa bacteriana en cultivos líquidos
Los métodos
mas comunes incluyen: b) El número de bacterias viables en un cultivo- usualmente se determina contando en número de colonias que crecen después de sembrar un volumen conocido sobre una placa (“conteo en placa” ó unidades formadoras de colonias). En ambos casos se debe graficar el logaritmo de la turbidez o el número de células vivas contra el tiempo y a este se le llama curva de crecimiento. El tiempo de generación se define como el tiempo requerido para duplicar la masa bacteriana del cultivo.
|
|||||||||||||||||||||||||||||||
|
RECURSOS DE LA WEB RUTA METABOLICA
Glicólisis |
METABOLISMO DE AZÚCARES (como un ejemplo de las rutas metabólicas). Glicolisis (Ruta de Embden, Meyerhof y Parnas [EMP] ) Esta es la ruta más común para el catabolismo de azúcares en las bacterias. (También se encuentra en la mayoría de las células de animales y vegetales). Una serie de procesos enzimáticos dan como resultado la conversión de azúcares para generar ATP (adenosin trifosfato) y NADH (nicotinamida adenina dinucleótido). La energía química necesaria para propósitos biosintéticos se almacena en compuestos formados de novo (ATP y NADH)
* referido al número de carbonos en la molécula. Existen rutas alternativas para el catabolismo de los azúcares a fin de producir energía almacenada en forma de ATP. Estos incluyen la ruta de la pentosa fosfato (derivada de la hexosa monofosfato) la cual se encuentra en la mayoría de las células vegetales y animales. El NADPH se produce usando esta ruta. Otra ruta, la de Entner Doudoroff, se encuentra en algunas bacterias. Respiración Anaeróbica La respiración anaerobia incluye la glicolisis y la fermentación. Durante los últimos estadios a este proceso, el NADH (generado durante la glicólisis) se reconvierte a NAD por pérdida de un hidrógeno. El hidrógeno se adiciona al piruvato y dependiendo de la especie bacteriana, se producen una variedad de productos finales metabólicos.
Respiración Aeróbica La respiración aeróbica incluye la glicolisis y el ciclo del ácido tricarboxílico (Ciclo de Krebs). El piruvato se degrada por completo hasta bióxido de carbono (C1) y en el proceso el NAD se convierte a NADH. Por lo tanto, en la fermentación aeróbica, el NADH se genera por dos rutas (glicolisis y ciclo de Krebs). La fosforilación oxidativa convierte el exceso de NADH a NAD y durante este proceso se genera más ATP (energía almacenada). Las ubiquinonas y los citocromos son componentes de la cadena de transporte de electrones, involucrada en este último proceso y la conversión de oxígeno hasta formar moléculas de agua viene a ser el paso final. Ciclo de Krebs (C4-C6 Compuestos intermediarios).
|
|||||||||||||||||||||||||||||||
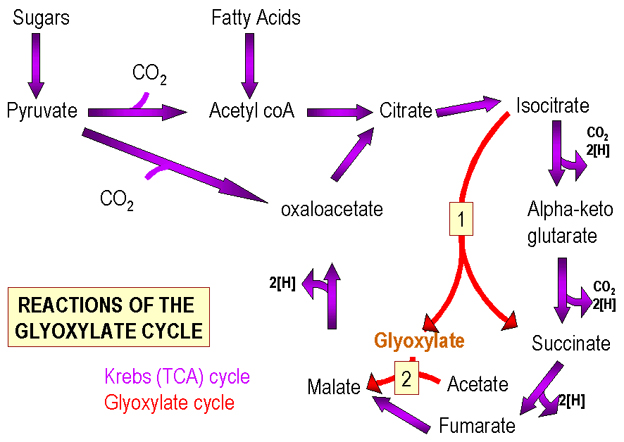 Figura 2. Los Ciclos de Krebs y Glioxilato
Figura 2. Los Ciclos de Krebs y Glioxilato |
Fosforilación Oxidativa
El Ciclo de Krebs (Figura 2) contiene intermediarios de 4 y 6 carbonos. El piruvato (C3) es la molécula que alimenta el ciclo de Krebs de tal manera que el número de intermediarios C4/C6 permanece igual o se incrementa. a) La pérdida de CO2 (C1) del piruvato para formar la acetil CoA, se ve seguida de su adición a un componente C4 del ciclo (oxalacetato) y se produce un componente C6 (ácido cítrico). Por lo tanto el número de moléculas C6 producidas iguala el número de moléculas C4 presentes inicialmente.
b) Mediante la adición del CO2 al piruvato se produce un compuesto de C4. En esta situación, se generan moléculas adiciónales de C4 (componentes del ciclo). .
Por
lo tanto si alguno de los componentes del ciclo, son removidos para usarse
en otras rutas biosintéticas, estos pueden ser repuestos por la vía de esta
reacción.
|
|||||||||||||||||||||||||||||||
|
RECURSOS DE LA WEB RUTAS METABOLICAS. |
||||||||||||||||||||||||||||||||
|
METABOLISMO DE LOS ACIDOS GRASOS
En lugar de ello, las bacterias utilizan el ciclo del glioxilato (Figura 2) (una modificación del Ciclo de Krebs) en el cual se saltan los pasos enzimáticos en los cuales dos moléculas de CO2 son removidas de los intermediarios de C6. El intermediario C6 se convierte en dos compuestos C4 (ambos componentes del ciclo). Por lo tanto por cada grupo acetilo (de los ácidos grasos) un intermediario adicional del ciclo se produce. La ruta del glioxalato generalmente no se encuentra en células animales, ya que se utilizan ácidos grasos preformados y presentes en los alimentos.
En resumen el del Ciclo de Krebs funciona para la producción de energía y biosíntesis de compuestos de carbono. Sin embargo, si los intermediarios del ciclo tienen que ser removidos para utilizarse en otras rutas metabólicas, siempre han de ser reemplazados. El proceso de reemplazamiento es diferente en el caso de la utilización de azúcares de aquel a partir de ácidos grasos.
|
|||||||||||||||||||||||||||||||
|
|
|
|||||||||||||||||||||||||||||||