| x | x | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DOENÇAS INFECCIOSAS | BACTERIOLOGIA | IMUNOLOGIA | MICOLOGIA | PARASITOLOGIA | VIROLOGIA | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| INGLÊS |
NUTRIÇÃO, CRESCIMENTO E METABOLISMO ENERGÉTICO
Dr Alvin Fox
Tradução: Dr. Paulo E. Moretti | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ESPANHOL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ALBANES | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PERSA | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ESCOLA DE MEDICINA DA UNIVERSIDADE DA CAROLINA DO SUL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dê a
sua opinião CONTATO |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
APRENDA PLUGADO |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Palavras-Chave Glicólise Fermentação Respiração anaeróbica Respiração aeróbica Anaeróbico facultativo Aeróbico obrigatório Anaeróbico obrigatório Anaeróbico aerotolerante Aeróbico microaerófilo Sideróforo Mesófilo Termófilo Psicrófilo Tempo de geração Ciclo do ácido tricarboxílico (CAT) ou Ciclo de Krebs Via do glioxilato ubiquinona
|
Os requerimentos para o crescimento das bactérias
incluem fontes de energia, carbono "orgânico" (por exemplo, açúcares e
ácidos graxos) e íons metálicos (por exemplo, ferro). Outros fatores
importantes são: temperatura ótima, pH ótimo e a presença (ou ausência) de
oxigênio. Requerimentos de oxigênio Bactérias aeróbicas obrigatórias requerem a presença de
oxigênio para poderem crescer; estas não realizam fermentação. Bactérias
anaeróbicas obrigatórias não efetuam a fosforilação oxidativa. Além disso,
elas são mortas pelo oxigênio. Essas bactérias não expressam certas
enzimas tais como a catalase [que quebra peróxido de hidrogênio,
H2O2, em água e oxigênio], a peroxidase [pela qual
NADH + H2O são convertidos a NAD e O2] e a
superóxido dismutase [pela qual o superóxido, O2-, é
convertido a H2O2]. Estas enzimas detoxificam os
radicais livres de peróxido e de oxigênio produzidos durante o metabolismo
na presença de oxigênio. Organismos anaeróbicos aerotolerantes são
bactérias que respiram anaerobicamente, mas que podem sobreviver na
presença de oxigênio. Bactérias anaeróbicas facultativas podem realizar
tanto fermentação quanto respiração. Na presença de oxigênio, a respiração
anaeróbica é geralmente desligada e esses microrganismos respiram
aerobicamente. Bactérias microaerófilas crescem bem em baixas
concentrações de oxigênio, mas são mortas em altas
concentrações. Requerimentos nutricionais Estes incluem fontes de carbono orgânico, nitrogênio,
fósforo, enxofre
e íons metálicos incluindo ferro. As
bactérias secretam pequenas moléculas que se ligam ao ferro (sideróforos,
por exemplo, enterobactina e micobactina). Os sideróforos (com ferro
ligado) são internalizados pela célula bacteriana após ligarem-se a
receptores específicos. O hospedeiro humano também tem proteínas
transportadoras de ferro (por exemplo, a transferrina). Desta forma,
bactérias que não competem eficazmente com o hospedeiro pelo ferro
disponível são pouco patogênicos. Temperatura As bactérias podem crescem em uma variedade de
temperaturas desde muito próximas do congelamento até próximas do ponto de
ebulição da água. Aquelas que crescem melhor em temperaturas medianas
neste espectro são denominadas mesófilas, as quais incluem todos os
patógenos e oportunistas humanos. Aquelas que crescem melhor em
temperaturas mais baixas ou mais altas são respectivamente denominadas
psicrófilas e termófilas. pH Muitas bactérias crescem melhor em pH neutro. Contudo,
certas bactérias podem sobreviver e mesmo crescer em condições de pH muito
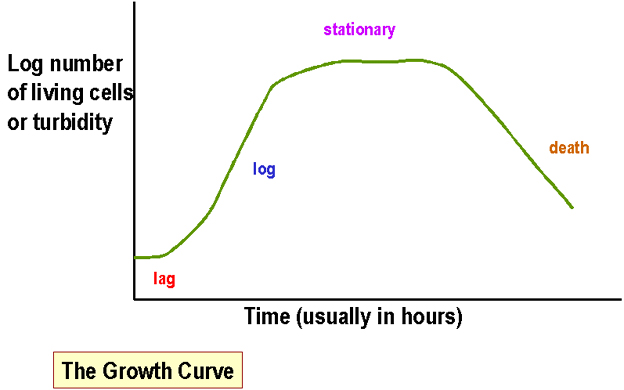
baixo ou muito alto. Medindo a massa de culturas bacterianas em meio líquido Os métodos mais comuns incluem medidas: a) da turbidez (a opacidade de uma cultura bacteriana em meio líquido - a medida das bactérias totais [vivas e mortas] - isto é usualmente quantificado com um espectrofotômetro). Figura 1. b) do número de bactérias viáveis em uma cultura -
usualmente obtido pela contagem do número de colônias que crescem em meio
sólido - em placa de Petri - após a semeadura de um volume conhecido da
cultura (contagem de unidades formadoras de colônia). Em cada um dos
métodos, gráficos construídos com o logarítmo das medidas de turbidez ou o
número de células viáveis versus unidades de tempo são denominados curvas
de crescimento. O tempo de geração é definido como o tempo requerido para
a massa bacteriana dobrar de tamanho. Metabolismo de açúcares (como exemplo de vias metabólicas) Glicólise (Via de Embden, Meyerhof and Parnas [EMP]) Esta é a via metabólica mais comum em bactérias para o metabolismo de açúcar (é também encontrada na maioria das células animais e vegetais). Uma série de processos enzimáticos resultam na conversão de açúcares em piruvato, gerando ATP e NADH (nicotinamida adenina dinucleotídio). A energia química necessária para propósitos biossintéticos é armazenada nos compostos formados (ATP e NADH).
* número de carbonos na molécula Há alternativas para essa via para catabolizar açúcares
com a finalidade de se produzir energia armazenada na forma de ATP. Estas
incluem a via da pentose fosfato (derivada da via hexose monofosfato) que
é encontrada na maioria das plantas e animais. O NADH é gerado através
dessa via. Outra via, a Entner Doudoroff, só é encontrada em algumas
bactérias. Respiração anaeróbica A respiração anaeróbica inclui a glicólise e a fermentação. Durante os últimos estágios deste processo, o NADH (gerado durante a glicólise) é convertido a NAD pela perda de um hidrogênio. O hidrogênio á adicionado ao piruvato e, dependendo da espécie da bactéria, são produzidos uma variedade de produtos metabólicos finais.
Respiração aeróbica A respiração aeróbica envolve a glicólise e ao ciclo do ácido tricarboxílico (ciclo de Krebs). O piruvato ó completamente degradado a dióxido de carbono (C1) e, no processo, o NAD é convertido a NADH. Desta forma, na fermentação aeróbica, o NADH é gerado a partir de duas rotas (glicólise e ciclo de Krebs). A oxidação fosforilativa converte o excesso de NADH a NAD e, no processo, mais ATP (energia armazenada) é produzido. As ubiquinonas e os citocromos são componentes da cadeia de transporte de elétrons envolvida neste último processo. A conversão de oxigênio a água é o passo final deste processo. Ciclo de Krebs (compostos intermediários C4-C6)
Fosforilação oxidativa
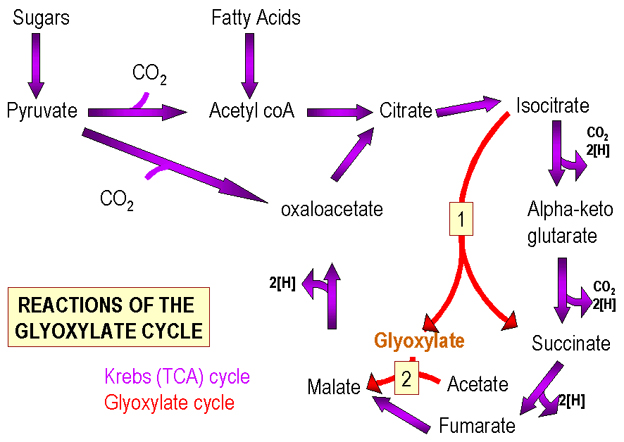
O ciclo de Krebs (Figura 2) contém intermediários de 4 e 6 carbonos. O piruvato (C3) supre o ciclo de Krebs de tal maneira que o número de intermediários de C4/C6 permanece o mesmo ou aumenta. a) a perda de CO2 (C1) do piruvato para formar acetil CoA, seguida de sua adição a um componente C4 do ciclo (oxaloacetato) produz um componente C6 (ácido cítrico). Assim, o número de moléculas de C6 produzidas se iguala ao número de moléculas de C4 inicialmente presentes.
b) pela adição de CO2 ao piruvato um composto C4 é produzido. Nesta circunstância, são formadas moléculas adicionais de C4 (um componente do ciclo).
Desta forma, se alguns dos componentes do ciclo são
removidos para uso em outras vias biossintéticas, estes podem ser repostos
por meio desta reação. Metabolismo de ácidos graxos Os ácidos são degradados a grupos acetil (C2) que suprem o ciclo de Krebs por sua adição a um intermediário C4 produzindo uma molécula C6. Durante o ciclo, o C2 adicionado é perdido como CO2 e C4 é produzido. Não ocorre aumento no número de moléculas intermediárias do ciclo. Assim, se ácidos graxos são a única fonte de carbono, nenhum intermediário do ciclo de Krebs pode ser removido sem que o ciclo se interrompa.
Ao invés, as bactérias utilizam o ciclo do glioxilato (Figura 2) (um ciclo de Krebs modificado) no qual não acontecem os passos enzimáticos em que duas moléculas de CO2 são removidas do C6 intermediário. Este último é convertido a dois compostos C4 (ambos componentes do ciclo). Desta forma, para cada grupo acetil (dos ácidos graxos) um ciclo intermediário pode ser produzido. Usualmente, a via do glioxilato não é encontrada em células animais uma vez que são utilizados ácidos graxos pré-formados presentes no alimentos.
Em resumo, o ciclo de Krebs funciona para produzir energia e compostos de carbono. Contudo, se os intermediários forem removidos para uso em outras vias metabólicas, estes devem ser repostos. O processo de reposição é diferente quando da utilização de açúcares ou ácidos graxos.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||