|
x |
x |
|
 |
 |
|
DOENÇAS
INFECCIOSAS |
BACTERIOLOGIA |
IMUNOLOGIA |
MICOLOGIA |
PARASITOLOGIA |
VIROLOGIA |
|
INGLÊS |
BACTERIOLOGIA – CAPÍTULO
QUATRO
MEMBRANA CELULAR, ESPOROS E BIOSSÍNTESE MACROMOLECULAR
Dr Alvin Fox
Emeritus Professor
University of South Carolina School of Medicine
Tradução: Dr.
Myres Hopkins
|
|
ALBANES |
|
ESPAÑOL |
|
PERSA |
|
ESCOLA DE MEDICINA DA
UNIVERSIDADE DA CAROLINA DO SUL |
|
Dê a sua opinião
CONTATO |
|
|
|
|
|
BUSCA |
|
E-MAIL
DR MYRES HOPKINS |
|

 |
|
APRENDA PLUGADO |
|
|
|
Palavras-Chave
Envoltório celular
Parede celular
Membrana Celular
Membrana externa
Peptidoglicano
Lipoproteína de Braun
Porinas
Lipopolissacarídeo (a “clássica” endotoxina)
Ácido teicóico
Ácido teicurônico
Ácido lipoteicóico
Ácido micólico
Undecaprenol (bactoprenol)
Endosporo
|
|
Nesta secção a estrutura dos envelopes celulares
Gram negativos, Gram positivos e envoltórios ácido-resistentes
será discutida. A composição e função de macromoléculas de
membrana especiais e sua biossíntese será descrita. Além disso
os endosporos, que são singulares em vários aspectos (inclusive
na estrutura do envoltório celular) serão discutidos.
Envoltório Celular
O envoltório cellular pode ser definido como a
membrana celular e parede celular mais uma membrana externa
quando presente. A parede celular consiste em uma camada de
peptidoglicano e estruturas a este ligadas. A maioria dos
envoltórios celulares se incluem em duas categorias principais
(Figura 1): Gram positivos e Gram negativos. Isso se baseia nas
características de se corar ou não pelo Gram que reflete
diferenças estruturais importantes entre esses dois grupos.
Outros tipos de parede celular são encontradas em algumas
espécies bacterianas (não Gram positivas ou Gram negativas).
O peptidoglicano é uma única macromolécula de
forma sacular, rica em ligações cruzadas que cerca a membrana
celular bacteriana conferindo-a rigidez. É enorme (bilhões em
peso molecular; compare com proteínas cujo peso molecular é na
ordem de milhões). Peptidoglicano consiste em um esqueleto de
glicano (polissacarídeo) que consiste em ácido N-acetil murâmico
e N-acetil glicosamina com cadeias laterais de peptídeos
contendo aminoácidos D- e L- e às vezes ácido diaminopimélico.
As cadeias laterais fazem ligações cruzadas por pontes
peptídicas. Essas pontes variam em estrutura entre espécies
bacterianas. Ácido murâmico, ácidos D-amino e ácido
diaminopimélico não são sintetizados por mamíferos. PG é
encontrado em totas as eubactérias exceto Clamidia e
Micoplasma. |
|
 Figura 1 Diagrama simplificado do envelope celular de bactéria Gram
positiva e Gram negativa
Figura 1 Diagrama simplificado do envelope celular de bactéria Gram
positiva e Gram negativa
|
|
Envoltório de Célula Gram Positiva
(Figura 2)
Covalentemente ligado ao espêsso peptidoglicano
está o ácido teicóico (seus esqueletos são usualmente polímeros
fosforados de ribitol ou glicerol) ou ácido teicurônico (ácido
glicurônico contendo polissacarídeos). Essas moléculas
negativamente carregadas estão supostamente envolvidas na
concentração de ions metálicos das redondezas. Ácidos teicóicos
podem também guiar enzimas autolíticas para os sítios de
digestão do peptidoglicano (autólise), uma das etapas da
biossíntese da parede celular. Algumas vêzes os polissacarídeos
neutros estão presentes. Ácido lipoteicóico, em muitas
bactérias, está geralmente associado com a membrana celular.
Outras vezes eles ocorrem nas fímbrias no lado de fora da
célula. Quando expresso no exterior da célula este ácido pode
estar envolvido na adesão a células epiteliais permitindo
colonização na garganta (ex. pelos streptococcus do grupo A). |
|
 Figura
2 Figura
2
As estruturas componentes do envoltório de célula Gram positiva
 Figura
3 Figura
3
As estruturas components do envoltório de célula Gram negativa
|
|
O envoltório de
Célula Gram negativa
(Figura 3)
Covalentemente ligado ao delgado peptidoglicano está a
lipoproteína de Braun que conecta a membrana externa à parede
celular. Assim como outras membranas, a membrana externa contém
proteínas e fosfolipídios. Diferentemente de outras membranas,
esta contém moléculas adicionais (lipopolissacarídeos). O
lipopolissacarídeo é importante para a bactéria uma vez que
ajuda a prover uma barreira de permeabilidade a substâncias
hidrofóbicas. O lipopolissacarídeo consiste em três regiões: um
antígeno O, uma parte central e uma região interna de lipídio A.
A parte central contém alguns açúcares (heptoses e
cetodeoxioctanato), não encontrado em outro lugar na natureza, e
lipídio A contém ácidos graxos β-hidroxílicos (incomuns na
natureza). A molécula
exibe atividade endotóxica. Porinas na membrana externa ajuda a
formar canais que permitem a passagem de nutrientes pequenos e
hidrófilos (tais como açúcares) através da membrana externa.
Bactérias ácido-resistentes e relacionadas
(micobactéria, nocardia e corinebactéria)
Os
envelopes celulares desses organismos são consideravelmente mais
complexos que de outras bactérias. Ácido micólico (ácidos graxos
de cadeia longa e ramificada) covalentemente conectado via um
polissacarídeo a peptidoglicano. Outros compostos de ácido
micólico e outros lipídeos complexos formam uma camada de
membrana cerosa espêssa do lado externo à camada de
peptidoglicano. |
|
 Figura 4 Estrutura do peptidoglicano de E. coli
Figura 4 Estrutura do peptidoglicano de E. coli
 Figura 5 Estrutura do peptidoglicano de S. aureus
Figura 5 Estrutura do peptidoglicano de S. aureus
|
|
Síntese de macromoléculas do envelope
celular
Peptidoglicano (Figuras 4 e 5): A subunidade precursora (muramil
pentapeptídeo ligado à uridina difosfato, UDP) é sintetizada no
citoplasma e passa para a membrana celular. A subunidade é
transferida enzimaticamente do nucleotídeo para um carreador
lipídico (undecaprenol/bactoprenol) e uma subunidade completa é
construída (dissacarídeo pentapeptídeo com ponte peptídica
anexada).
As
subunidades completas são então exportadas para a parede
celular. Após a liberação do monômero o undecaprenol é
recirculado na membrana celular e reutilizado. O esqueleto de
glicano da parede celular existente é degradado enzimaticamente
(por autolisinas) para permitir a inserção da nova subunidade
sintetizada. Se essas enzimas forem superativas, a parede
celular se degrada e a elevada pressão osmótica da célula rompe
a membrana citoplasmática matando a célula (“autólise”). A
ligação cruzada da cadeia lateral do peptídeo da subunidade
inserida à cadeia existente ocorre então enzimaticamente
(proteínas ligadoras a penicilina). Subunidades completas de
ácido teicóico e teicurônico são também sintetizadas na membrana
celular (em carreadores lipídicos) antes do transporte e
inserção à parede celular existente. |
|
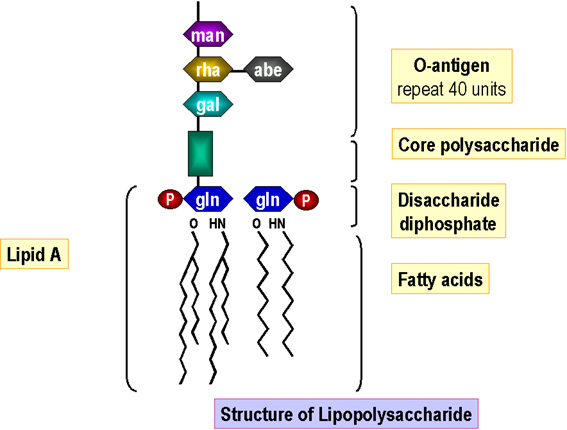
 Figura
6. Estrutura de lipopolissacarídeo
Figura
6. Estrutura de lipopolissacarídeo |
|
Lipopolissacarídeo
O
lipídeo A é montado na membrana celular e a base de açúcares
anexada sequencialmente. Subunidades de antígeno O são
sintetizadas independentemente (em lipídeo carreador como na
síntese do peptidoglicano). O antígeno O sintetizado é então
anexado ao centro de lipídeo A (gerando lipopolissacarídeo) na
membrana celular antes da passagem/inserção na membrana externa
(Figura 6). |
|
 Figura 7. Etapas da formação do esporo
Figura 7. Etapas da formação do esporo
|
|
Endosporos
Essas células bacterianas Gram positivas modificadas têm um
envoltório celular incomum que contém uma membrana celular e uma
membrana externa. A camada de peptidoglicano é menos dotada de
ligações cruzadas que na maioria das células bacterianas e
contém uma forma desidratada de ácido murâmico. O peptidoglicano
do esporo é referido como um cortex e é encontrado entre as duas
membranas. Uma capa consistindo de queratina com alto teor de
ligações cruzadas é encontrada ao redor do lado externo da
célula. O esporo bacteriano é altamente resistente a agentes
químicos por causa dessa capa.
Normalmente na replicação bacteriana, as células dividem, um
septo se forma dividindo a célula mãe em duas células-filhas de
tamanho aproximadamente igual. Quando a esporulação ocorre, a
divisão celular é desigual e a célula-mãe que é maior envolve a
célula-filha. A membrana celular da célula-filha constitui a
membrana interna do esporo e a membrana celular da célula-mãe
forma a membrana externa (Figura 7 e 8). |
|
|
FILME
Formação do Endosporo
Requer Flash
© Joseph J. Gauthier and Jane Bynum
University of Alabama - Birmingham
Birmingham, AL and
The MicrobeLibrary
|
|
|
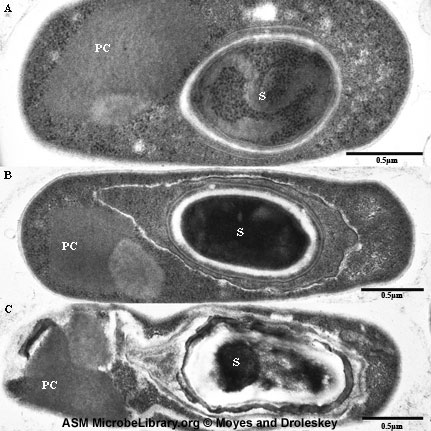
 Figura 8A
Bacillus mostrando células vegetativas e células contendo endosporos.
Coloração pelo Gram
Figura 8A
Bacillus mostrando células vegetativas e células contendo endosporos.
Coloração pelo Gram
© Christopher J. Woolverton, Kent State University Kent, Ohio e
The MicrobeLibrary
 Figura 8B
Bacillus thuringiensis com o corpo parasporal ao lado do endosporo em
formação. O corpo parasporal (ou cristal parasporal) age como bio-pesticida
contra o estágio larval de mais de 100 espécies de mariposas.
Figura 8B
Bacillus thuringiensis com o corpo parasporal ao lado do endosporo em
formação. O corpo parasporal (ou cristal parasporal) age como bio-pesticida
contra o estágio larval de mais de 100 espécies de mariposas.
© Rita Moyes, Robert Droleskey and
The MicrobeLibrary
|
|
|
 Retorne à Seção de Bacteriologia de Microbiologia e
Imunologia On-line
Retorne à Seção de Bacteriologia de Microbiologia e
Imunologia On-line
Esta página foi traduzida do original em inglês por Myres MTR Hopkins, PhD
em Ciências (Genética – Universidade de São Paulo) e é mantida por Richard
Hunt
Por favor, relate quaisquer problemas para
mmtr5@hotmail.com
|
 Figura 1 Diagrama simplificado do envelope celular de bactéria Gram
positiva e Gram negativa
Figura 1 Diagrama simplificado do envelope celular de bactéria Gram
positiva e Gram negativa Figura
2
Figura
2  Figura 4 Estrutura do peptidoglicano de E. coli
Figura 4 Estrutura do peptidoglicano de E. coli Figura
6. Estrutura de lipopolissacarídeo
Figura
6. Estrutura de lipopolissacarídeo Figura 7. Etapas da formação do esporo
Figura 7. Etapas da formação do esporo


{kind=link}