|
x |
x |
|
 |
 |
|
INFECTIOUS
DISEASE |
BAKTERIYOLOJİ |
İMMÜNOLOJİ |
MYCOLOGY |
PARASITOLOGY |
VİROLOJİ |
|
ENGLISH |
VIROLOJİ - BÖLÜM YİRMİBEŞ
KORONA VIRUSLAR, SOĞUK ALGINLIĞI VE SARS
Dr Richard Hunt
Professor
Department of Pathology, Microbiology and Immunology
University of South Carolina School of Medicine
Çeviren
Prof. Dr. Selçuk Kaya
İzmir Katip Çelebi Üniversitesi, Tıp Fakültesi
|
|
ALBANIAN |
|
Español |
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|

 |
|
Logo image © Jeffrey
Nelson, Rush University, Chicago, Illinois and
The MicrobeLibrary |

 Şekil 1 Koronaviruslar mikroskop altında
bakıldığında bir halo veya taç benzeri (corona) görünüme sahip
virusların bir grubu
Şekil 1 Koronaviruslar mikroskop altında
bakıldığında bir halo veya taç benzeri (corona) görünüme sahip
virusların bir grubu
CDC/Dr. Fred Murphy (top) CDC/Dr. Erskine Palmer (bottom)
|
Coronavirüsler 100 nm büyüklüğünde olup, en büyük pozitif polariteli RNA
virüsleridir (böylece tüm RNA virüsleri içinde en büyük genoma sahip
virüslerdir). İnsan ve hayvanları enfekte ederek respiratuar ve enterik
hastalıklara neden olurlar. Nidovirales ailesinden, Torovirüs ve
Arterivirüslerle aynı gruptadır, bir dizi mRNA ile ortak 3’sonlanmasını
oluşturur. Coronavirüsler ve Torovirüsler (birlikte Coronaviridea’yı
oluştururlar) helikal nükleokapsitlidir; Arterivirüsler ikozahedral
nükleokapsitlidir. Coronavirüsler intrasellüler membrandan köken alan
zarfa sahiptir (plazma membranı değil). İsminden anlaşılacağı üzere
elektron mikroskopunda yüzeyden çıkıntı yapan dikenleri vardır (geniş
glikoprotein nedeniyle) (corona=taç) (Şekil 1,2).
Bu bölümde sadece Coronavirüsler tartışılacaktır çünkü respiratuar
hastalıklardan ve soğuk algınlığının üçte birinden sorumlu olmaları ve
SARS (severe acute respiratory syndrome)’da etken olmaları nedeniyle
özel bir öneme sahiptir.
|

Şekil 2 Torovirus
© Queen's University, Belfast
 Şekil 3
Şekil 3
Koronavirus yapısı.
Adapted from Lai and Homes. In Fields' Virology.
Lippencott
|
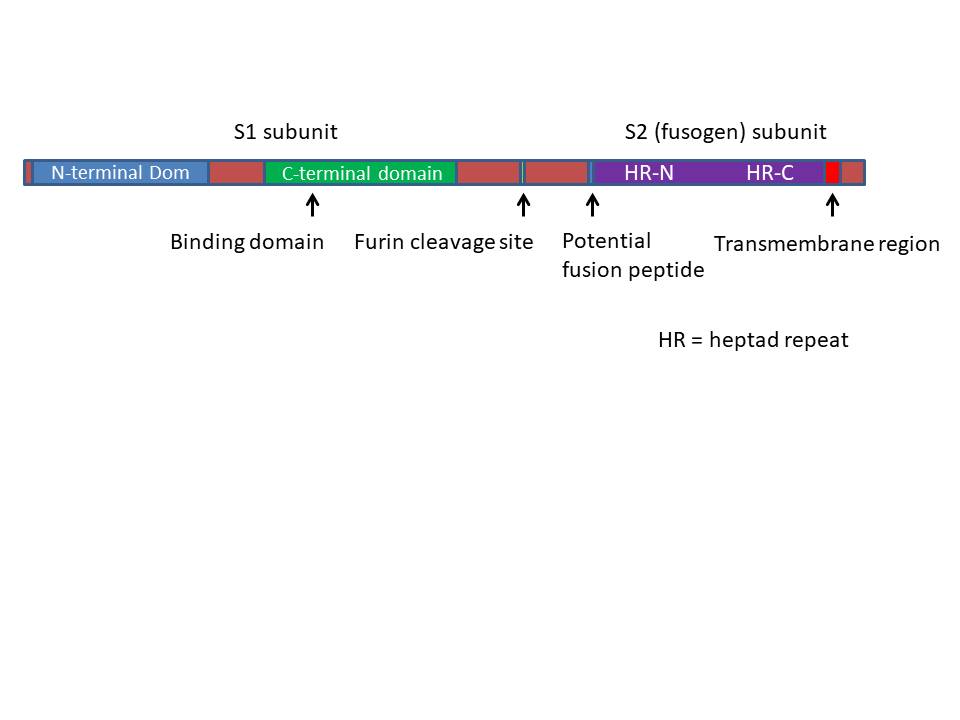
PROTEINLER
S (spike=diken) protein (150k)
S proteini 3 bölgesi olan bir transmembran proteindir: büyük
eksternal bölge, transmembran sekansı, küçük internal bölge. Dış bölgesi
(N-terminal) globuler formda katlanır ve EM’da diken yapıları
gözlenebilir. Bu bölge virüse antijenik özelliklerini kazandırır ve
hücre yüzey reseptörlerine bağlanabilmesi için reseptör oluşturur. Dış
bölgenin iç kısmı sarmal yapıda olup, yedili tekrarlar içerir. Lipit
çift tabakada proteinleri sabitlemek için yağ asiti molekülü
bulunmaktadır. Dış bölgenin iç kısmı sap benzeri bir yapıda olup trimer
yapı oluşturmak için diğer S proteinleriyle ilişki içindedir. Bazı
Coronavirüslerde dış bölge bölünmüş haldedir fakat bu iki parçalı
glikoprotein yapı iyonik bağlar sayesinde birbirleriyle etkileşim
içindedir (HIV gp120 ve gp41’de olduğu gibi). S proteinin iç kısmı konak
hücreye bağlanma aşamasında, membran füzyonundan sorumludur. İlginç
olarak, S protein immünoglobulinlerin Fc-gama reseptörlerine benzer bir
bölgeye sahiptir; virüse bu proteinlerle kaplanma ve immün yanıttan
kaçma şansı verir (herpes virüs de benzer stratejiye sahiptir). S
protein konak hücre yüzeyinde sialik asite bağlanabilir; böylece virüs
hemaglütinasyon yeteneğine sahip olmuş olur. S proteinine karşı
antikorlar nötralizan özelliktedir.
HE proteini (65kD)
Sadece bazı Coronavirüsler hemaglütinin-esteraz proteinine sahiptir.
Virüs yüzeyinde S protein dikenlerinden daha kısa dikensi yapıları
vardır. Dimer yapıdadır ve replikasyon için elzem değildir. Bu protein
ayrıca sialik asite bağlanabilir. Hemaglütinin-esteraz proteininin
esteraz aktivitesi sialik asiti şeker zincirinden ayırabilir, böylece
replike olduktan sonra hücreden çıkmaya yardımcı olur.
Hemaglütinin-esteraz proteinine karşı oluşmuş antikorlar da virüsü
nötralize edebilir.
M (membran) proteini
Diğer bir membrana bağlı glikoprotein olan M (membran) proteini,
internal protein olup sadece küçük bir eksternal N-terminal domain
içerir. M proteini viral membranı üç kez geçmektedir. Bu protein,
nükleokapsitin golgi cisimciği gibi internal membranlara bağlanmasını
kolaylaştırır ve diğer glikoproteinler gibi plazma membranında bulunmaz.
E (zarf) proteini (9-12kD)
Bu küçük protein de viral membranda bulunur. Bu protein enfekte
hücrelerde nükleus çevresinde ve hücre yüzeyinde bulunur.
N (nukleokapsid) protein (60kD)
Nükleokapsit proteini lider sekans ve viral membranın iç yüzündeki M
proteini aracılığıyla genomik RNA’ya bağlanır. N proteini
fosforillenmiştir.
Diğer RNA virüslerinin aksine Coronavirüs bünyesinde RNA polimeraz
enzimini içermez; bunun yerine enfeksiyon oluştuktan sonra pozitif
polariteli genomik RNA mRNA olarak kullanılarak bu enzim oluşturulur.
Pol geninin genomun 5’ ucunda bulunması sayesinde bu mümkün olmaktadır.
|
| |
KONAK HÜCREYE VIRUSUN TUTUNMASI
Daha önce belirtildiği üzere S proteini majör bağlantı proteini olup,
sialik asite bağlanır. HE proteini de sialik asite bağlanır. Sialik asit
tüm hücre yüzeylerinde bulunur ve Coronavirüsler belli dokulara tropizm
gösterirler; bu yüzden bağlanma daha komplike mekanizmalarla olmaktadır.
Dahası bazı Coronavirüsler tüm sialik asitlere bağlanmaz. S protein
başka daha spesifik reseptörlere bağlanabilir. Fare hepatit virüsünün
reseptörü immünoglbulin süperailesinin üyesi olup, bu proteine karşı
antikor virüs bağlanmasını engellemektedir. İnsan respiratuar
Coronavirüsleri de dahil olmak üzere diğer Coronavirüsler reseptör
olarak membrana bağlı bir metalloproteinazı (aminopeptidaz N)
kullanırlar.
|
| |
PENETRASYON Sitoplazmada
viral replikasyon için viral membran ile hücre membranının füzyonu
öncelikli olup, asidik endozom veya plazma membranında gerçekleşebilir.
|
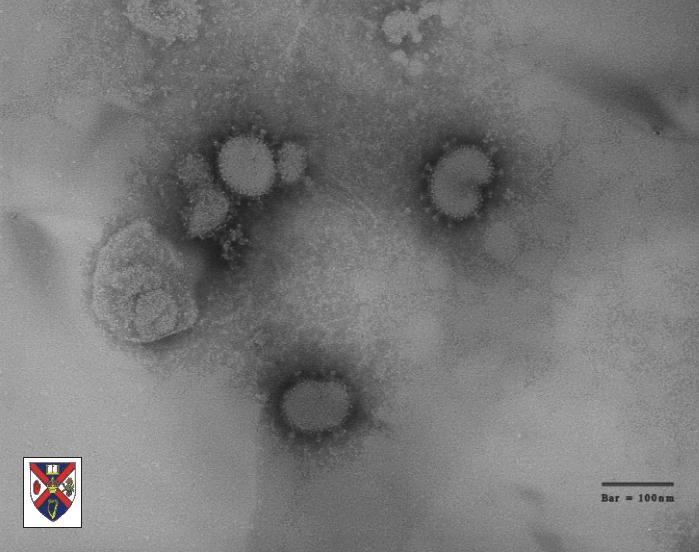
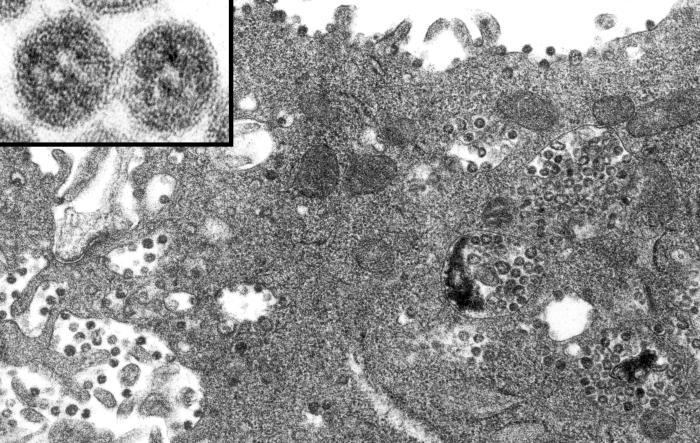
 Şekil
4 Şekil
4
Sitoplazmik zara bağlı vakuol ve granüllü endoplazmik retikulum
sisternalinde koronavirus. Enfekte Vero E6 hücresinin ince kesit
elektron mikroskobu, şiddetli akut solunum yolu sendromu (SARS) nedeni
olarak şüphelenen koronavirus parçacıklarını gösterir.
CDC/C.S. Goldsmith/T.G. Ksiazek/ S.R. Zaki |
VIRUS BİRLEŞMESİ
Sadece pozitif polariteli genomik RNA üzerinde bulunan, genomun 3’ ucuna
yakın bölgesinde 61 nükleotidlik sekans bölgesi bulunmaktadır. Bu bölge
nükleokapsiti oluşturmak üzere N protein ile ilişki içindedir.
Nükleokapsit ileintrasellüler membranın (endoplazmik retikulum, golgi
cisimciği ve bu ikisi arasındaki bölge, muhtemelen cis-Golgi ağı)
sitoplazmik yüzünde bulunan M proteini bağlantı halindedir. M proteini
plazma membranınınn sitoplazmik yüzünde değildir. Bunun oluşabilmesi
için N proteininin RNA ile iletişim halinde olması gerekmektedir. Dahası,
E proteini tomurcuklanma işlemi için membran eğimini değiştiren
nükleokapsitin tomurcuklanması için gerekmektedir. M proteinine bağlı E
proteinidir bu. E proteini olgun virüste düşük miktarda bulunur,
böylelikle toplanmanın başlayabilmesi için yapı proteini işlevi görür.
Ayrıca S ve HE proteini tomurcuklanan bölgede lipit çift katmanı içinde
M proteini ile bağlantı halindedir. Virüs golgi cisimciğinden geçerken
S-M ve HE-M potein kompleksleri birleşir ve şeker zincirleri oluşturulur.
Virüs morfolojik olarak golgi cisimciğinde olgunlaşır ve sitoplazmada
sonradan membranla kaynaşacak olan membrana bağlı veziküllerde birikir (Şekil
4).
|
| |
GENOM Coronavirüsler
tek zincirli oldukça büyük bir genoma sahiptir, aslında tüm RNA
virüsleri arasında en büyük genoma sahip olanıdır. Genom pozitif
polaritelidir (yani mRNA ile aynı) ve segmentsizdir (ortomiksoviruslar).
Genomik RNA poliadenilli ve başlıklı olup 27-32 kB büyüklüğündedir. Bu
büyük genom yapısında RNA polimerazın proof reading aktivitesi yoktur ve
bu durum Coronavirüslerde yüksek mutasyon düzeyine neden olur. SARS
virüs dahil birçok Coronavirüs sekanslandı. Gen ürünleri daima aynıdır.
5’ ucunda polimeraz (pol) bulunur ve bunu tüm Coronavirüslerde bulunan
dört yapısal protein izler:
- Spike protein (S), bu şekilde adlandırılmasının nedeni olarak
virüs yüzeyinden çıkıntı yapması gösterilebilir.
- Envelop protein (E)
- Membran protein (M), konak hücrenin intrasellüler membranıyla
bütünleşir.
- Nükleokapsid protein (N)
Bazı coronavirüsler hemaglütinin-esteraz (HE) adı verilen, pol ve S
genleri arasında bulunan ve paramiksoviruslardan kazanılmış olan bir gen
taşır. Ayrıca birçok coronavirüste open reading frames (ORF) denen çok
iyi korunmamış alanlar vardır. Bu genler fonksiyonları tam olarak
bilinemeyen proteinler kodlar. Genomik RNA’daki protein kodlayan genlere
ek olarak tüm coronavirüsler her bi genin 5’ ucunda intergenik sekanslar
olarak adlandırılan 7 baz sekansı içerir. Eğer intergenik sekans
mutasyona uğrarsa sub-genomik mRNA bu alandan başlamak üzere kodlanmaz.
|
 Şekil 5
Şekil 5
Koronaviruslarının messenger RNA'ları. Ortak bir 3 'ucu ile RNA'lar iç
içe geçmiş dizi oluşturulur. Polimeraz için mRNA (pol) genomik RNA ile
aynı uzunluktadır. hepsi ortak bir lider diziye sahip olmasına rağmen,
geri kalan 5 'ucunda kesilir |
REPLIKASYON Enfekte
hücrede genomik RNA’nın kaderi, viral partiküle katılmaktır. Bu
moleküller büyük ihtimalle, mRNA’nın aksine sürekli sentez edilmektedir,
mRNA sentezinde kullanılmak üzere lider sekansın parça parça
üretildiğine dair kanıtlar mevcuttur (aşağı bakınız).
Messenger RNA
Tüm coronavirüsler yaygın 3’ ucu içerirken, 5’ sonlanmasından
mahrumdur (gen 1 proteinini (polimeraz) kodlayan mRNA hariç) (Şekil
5). Genomik RNA gibi sub-genomik mRNA’lar da başlıklı ve
poliadenillidir. Sadece tek bir protein her bir subgenomik mRNA
tarafından kodlanır, bu protein 5’ ORF bölgesinde kodlanır. Hatta en
küçüğü hariç tüm bu moleküller birden fazla sekansa sahiptir. Her
mRNA 5’ ucunda 70 baz uzunluğunda yaygın lider sekansa sahiptir.
Genomik RNA da 5’ ucunda bu bölgeye sahiptir fakat intergenik
sekansları, lider zincir ile benzerdir.
Genomik RNA sitoplazmaya girdiğinde tamamlayıcı negatif zincir
sentezlenir. Ardından tekrar genomik pozitif zincire ve subgenomik
mRNA ya dönüştürülür. Hücrede ayrıca subgenomik negatif zincir RNA
bulunur fakat bunlar daima çift zincir halinde bulunurlar.
Subgenomik mRNA ların lider sekanslardan nasıl yapıldığı tam olarak
bilinememekle birlikte bu konuda birkaç olasılık söz konusudur.
Örneğin, negatif zincirin lider sakansı 70 bazlık pozitif zincire
kopyalanıyor olabilir. Lider sekans ardından genomik zincire ayrışır
ve kalıp üzerindeki intergenik sekanslardan birini tanıyabilir. Bu
genomik RNA’nın sonuna kadar ilk pozitif zincir sentezi olabilir.
mRNA sentezindeki bu kesikli modeli destekleyen gözlem şudur; eğer
bir intergenik sekans dış müdahale ile genoma yerleştirilirse, bu
noktadan itibaren lider sekansı içeren yeni bir mRNA başlamış olur.
|
| |
Polimeraz (replikaz)
Genomun 5’ ucundaki ilk gen replikaz veya RNA polimerazı kodlayan
gendir. 20 Kb’lık uzunluğuyla genomun yarısından daha fazlasını
kaplamaktadır. Sekanslama gösterdi ki bu gen aslında iki protein
kodlayan bir alandır, birbiri üzerine binebilir ve farklı
ORF’lardadır. Fakat bu iki sekans, ribozom ikinci sekans bölgesine
geldiğinde ribozomal kayma ile poliprotein adındaki tek bir proteini
kodlamaktadır. Bu dev poliprotein yapıldığında, ilk formunun bir
parçası olan proteazlar tarafından parçalanır. Bu proteinlerden biri
olan RNA polimeraz salınmış olur.
Mutasyon ve rekombinasyon
Coronavirüsler viral replikaz/polimeraz aracılığıyla kodlanan
geniş bir RNA genomuna sahiptir. RNA polimeraz proof reading
aktivitesine sahip olmadığı için 10bin nükleotidde bir hata payına
sahiptir. Ortalama bir coronavirüsün 30kB’lık bir büyüklükte olduğu
düşünülürse, progeny virüste oldukça fazla sayıda mutasyon olacaktır.
Coronavirüslerde ayrıca birçok delesyon mutasyonu gerçekleşir.
Segmentsiz RNA virüslerinde tipik olmamakla birlikte
coronavirüslerde oldukça fazla bir şekilde rekombinasyon olmaktadır.
Bu durum RNA replikasyonunun kesikli bir şekilde yapılması ve önce
lider sekansın yapılması ardından polimerazın bir başka zincire
atlamasındandır. Bu yüksek rekombinasyon sıklığı virüsün hızlı
evrimleşmesine ve yeni suşların oluşmasına neden olur.
|
| |
SOĞUK ALGINLIĞI VE
KORONAVIRUSLARIN DİĞER HASTALIKLARI
Patogenez
Coronavirüsler birçok hayvanda respiratuar ve enterik
hastalıklara neden olur. İnsanlarda virüsün majör replike olduğu
alan respiratuar epitel hücreleridir ve soğuk algınlığının üçte biri
coronavirüsler tarafından oluşturulur. Rhinovirüsün sebep olduğu
soğuk algınlığıyla benzer semptomlara neden olur (burun akıntısı,
boğaz ağrısı, öksürük, başağrısı, ateş, üşüme vb.) ve inkübasyon
süresi yaklaşık 3 gündür. Viral yayılım birçoklarında immün sistem
tarafından sınırlandırılır fakat immünitesi kısa sürelidir.
Zarflı rhinovirüslerin aksine coronavirüsler daha kararsız
yapıdadır. Hapşırma gibi aerosol oluşturan nasal sekresyonlar
aracılığıyla yayılır. Virus bağırsak epitel hücrelerini enfekte
ederek diyareye neden olur. Bu durum yenidoğanlarda görülebilir;
fakat birçok hayvan yavrusu için daha sıktır ve fatal olabilir.
Coronavirüs enfeksiyonları genellikle lokalize kalsa da yayılabilir.
İnsanlarda, orta kulak iltihabına, immünsuprese hastalarda pnömoniye
ve myokardite sebep olabilir fakat hayvanlarda sistemik enfeksiyon
çok daha ciddi seyreder (örn; kedi enfeksiyöz periodontiti).
İlginç olarak laboratuvar koşullarında nöral hücreleri enfekte
edebilen coronavirüsler rodentlerde multiple sklerozise benzer
hastalık oluşturabilir, böylelikle insanlarda da hastalık
yapabildiğine dair kanıt teşkil etmektedir. Rodentlerde
demyelinizasyon ile karakterize multiple sklerozisin, S protein ile
ilişkili olup bu proteine karşı oluşmuş immün yanıtın, moleküler
benzerlik neticesinde myeline karşı oluştuğu öne sürülmüştür.
Hastaların beyninde virüs izole edilmiş olmasına rağmen, multiple
skleroz ile olan ilişkisi henüz kanıtlanamamıştır.
|
| |
Epidemioloji
Birçok kişide anti-coronavirüs antikoru bulunmasına rağmen,
reenfeksiyon sıktır. Bu durum popülasyonda virüsün birçok
serotipinin olduğunu göstermektedir. İnsanları infekte eden bu
viruslar için hayvan rezervuarı gösterilememiştir.
Diğer birçok respiratuar enfeksiyonda olduğu gibi coronavirüs
kaynaklı soğuk algınlığı da sıkı temas nedeniyle kış aylarında daha
yoğundur. Birkaç yılda bir virüs tipine bağlı olarak büyük salgınlar
meydana gelir.
Tanı
Birçok coronavirüs enfeksiyonu tespit edilemez ve kendi kendini
sınırlar. Tanı immünoelektron mikroskopisi ve serolojiyle konur.
Coronavirüs enfeksiyonu için rutin kullanımda anti-viral ajan
bulunmamaktadır fakat semptomları hafifletmek için anti-viral terapi
uygulanabilir.
|
|
|
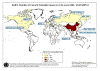 Şekil 6A
Şekil 6A
Muhtemel SARS vakaları haritası. 02 Haziran, 2003
WHO
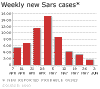 Şekil 6B
Şekil 6B
Haftalık yeni SARS vakaları.
WHO/BBC
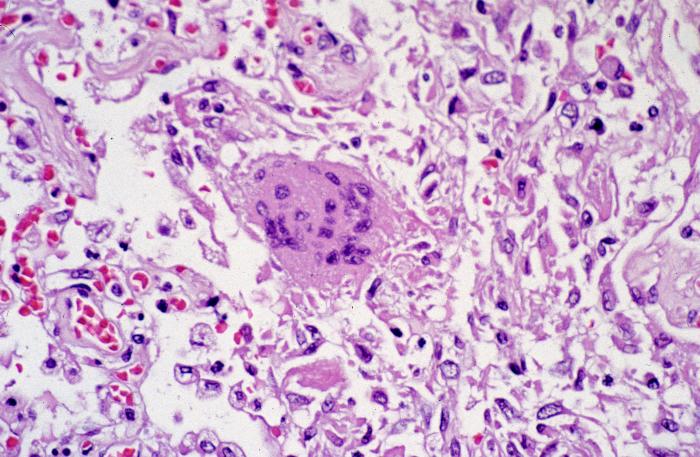
 Şekil 7
Şekil 7
Patolojik cytoarchitectural değişiklikler, göze çarpmayan viral
inklüzyonlar ile çok çekirdekli dev hücreler kadar diffüz alveoler
hasarın göstergesidir.
CDC/Dr. Sherif Zaki
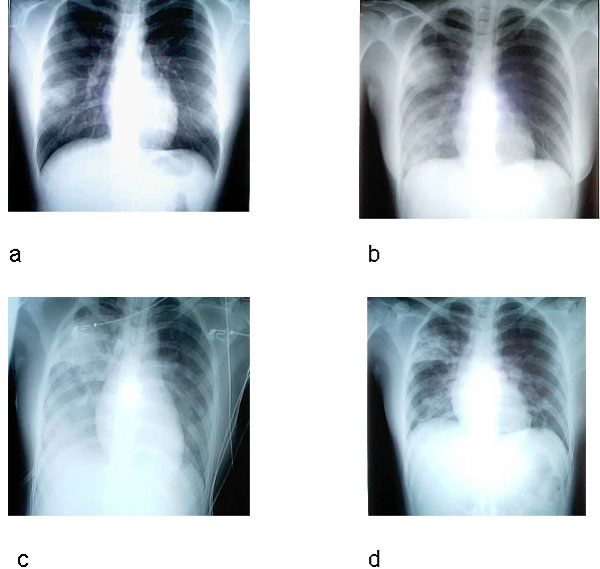
 Şekil 8
Şekil 8
Şiddetli akut solunum yolu sendromlu (SARS) endeks hastanın akciğer
grafileri. a, semptomların 5. günü; b gün 10; c, gün 13; d gün 15.
Li-Yang Hsu, Cheng-Chuan Lee, Justin A. Green, Brenda Ang,
Nicholas I. Paton, Lawrence Lee, Jorge S. Villacian, Poh-Lian Lim, Arul
Earnest, and Yee-Sin Leo - Tan Tock Seng Hospital, Tan Tock Seng,
Singapore. Emerging and Infectious Diseases
|
ŞİDDETLİ AKUT SOLUNUM YOLU SENDROMU (SARS)
2002’lerin sonlarına doğru güney Çin’de Guangdong blögesinde yeni bir
sendrom ortaya çıktı. Akut şiddetli respiratuar senndrom (SARS) olarak
adlandırıldı. Şu anda Asya, Kuzey Amerika ve Avrupa’dan (Şekil 6A) rapor
edilen hastalık başağrısı, genel kırgınlık ve ağrının eşlik ettiği 38C
üzeri ateş ile karakterizedir. Aslında respiratuar semptomlarbaşlangıçta
orta düzeyde iken birkaç gün veya bir hafta içinde hasta kuru öksürük ve
solunum sıkıntısı (dispne) ile gelebilir. Vakaların %3-30’unda solunum
sıkıntısı ölüme neden olmaktadır. Laboratuar olarak lenfosit sayısında
düşme ve karaciğer hasarını gösteren aminotransferaz aktivitesinde artış
görülür.
İlk SARS salgını 2003 yılının nisan ayında başladı ve haziran ayında
etkisi azalmaya başladı. Bu süre içinde dünya çapında 8000 vaka ve 775
ölüm meydana geldi. Dahası milyarlarca dolar ekonomik kayba neden oldu.
Virüs Vero E6 maymun hücre kültüründe geliştirildi ve bu yeni
coronavirüs (SARS-coV) hastalıkla ilişkili bulundu. 29727 bazlık ve 11
ORF’e sahip bir genomdur. Sekans yapısı, bu virüsü yeni bir coronavirüs
grubu üyesi yapmaya yetecek kadar farklıdır. Genom yapısı diğer
coronavirüslerle oldukça benzerdir (5’ replikaz, spike (S), zarf(E),
membran (M), nükleokapsit (N)-3’ ve her iki terminalde bulunan kısa
kodlanmayan bölgeler ). Replikaz geni genomun 5’ ucundan üçte ikisini
kaplar ve diğer coronavirüsler gibi iki ORF bölgesi üstüstedir. Ayrıca
Pol poliproteininden bir proteaz kodlar. Diğer coronavirüslerde
bulunmayan 9 ORF bölgesi vardır ve SARS virüsüne özel bazı proteinler
sentezliyor olabilir. Antikor testleri kullanılarak SARS-coronavirüs,
dünya genelinde SARS hastalığıyla ilişkili bulunmuştur.
Tanı
Hastalık Kontrol Merkezi göğüs X-ray,
pulse-oksimetri, kan kültürü, balgam kültürü ve Gram boyaması ve
özellikle RSV, İnfluenza A ve B olmak üzere viral respiratuar
patojenler açısından test edilmesini önermektedir. Legionella ve
pnömokokal üriner antijen testleri için örnekler alınmalı. SARS
olduğundan şüphelenilen kişiler izole edilmeli ve karantina altına
alınmalıdır.
Tedavi
Semptomatik tedaviden başka SARS için
görüş birliğine varılmış bir tedavi protokolü yoktur. Yeni ilaçlar
geliştirilmeye çalışılmakta ve proteaz fonksiyonunu baskılayan
ilaçlar üzerinde yoğun çalışmalar mevcuttur. SARS virüsü veya diğer
insan coronavirüslerine karşı aşı yoktur. Bazı coronavirüs suşlarına
karşı veteriner aşı programlarında orta derecede başarı sağlanmıştır.
Canlı viral aşı için, virüsün antijenik shifte uğraması ve tahmin
edilemeyen etkiler majör problemi oluşturmaktadır.
YENİ BİR
KORONAVIRUS – ORTA ASYA RESPIRATUAR SENDROM KORONAVIRUS
2012’de Ortadoğu’da özellikle Suudi Arabistan’da yeni bir coronavirüs
kaynaklı hastalık ortaya çıkmıştır. Başlangıçta tüm hastalar ya
Ortadoğu’da yaşayan ya da o bölgeyi ziyaret eden hatta sonradan
Avrupa’ya göç etmiş kişilerde görülüyordu. İlk enfeksiyon dalgasından
sonra virüs, insandan insana yakın temas ile yayılmaya başladı. Hastalar
fatalite oranının %50’nin üzerinde olduğu pnömoni ve böbrek yetmezliğine
girmeye başladı. Fakat bu yüksek fatalite düzeyi daha az virülen
vakaların tanısını önlüyor olabilir. Bu virüs önceleri Yeni Corona Virüs
diye adlandırılmış, daha sonra Ortadoğu Respiratuar Sendrom Coronavirüs
(MERS-CoV) diye adlandırılmış ve SARS coronavirüsten ayrıştırılmıştır.
İnterferonα2b ve ribavirin ile tadavi ediliyor.
Tekrardan bu yeni coronavirüsün kaynağı yarasalar ve özellikle
pipistrelle yarasalarıdır ve filogenetik analiz araştırmalarına göre
insanlara 2011 yılında geçmiştir.
|
|
|
| |
|
|
|
 Mikrobiyoloji ve İmmünoloji On-line, Viroloji Bölümüne Dönünüz
Mikrobiyoloji ve İmmünoloji On-line, Viroloji Bölümüne Dönünüz
This page last changed on
Wednesday, November 23, 2016
Page maintained by
Richard Hunt
Please report any problems to
Richard Hunt
|

 Şekil
4
Şekil
4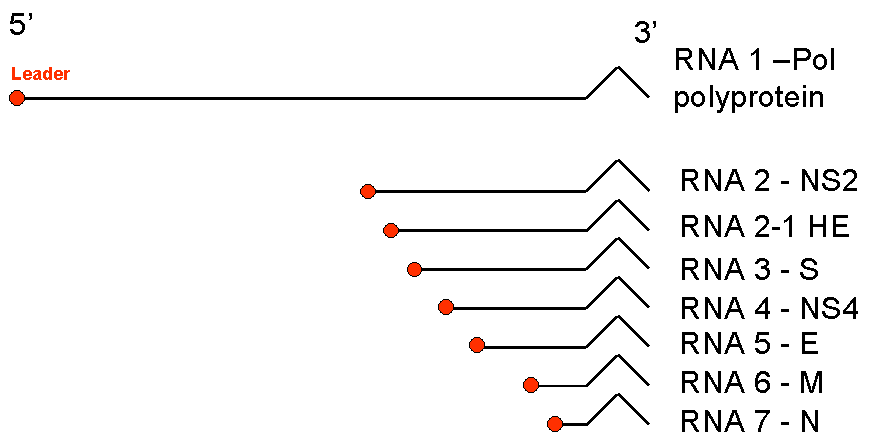 Şekil 5
Şekil 5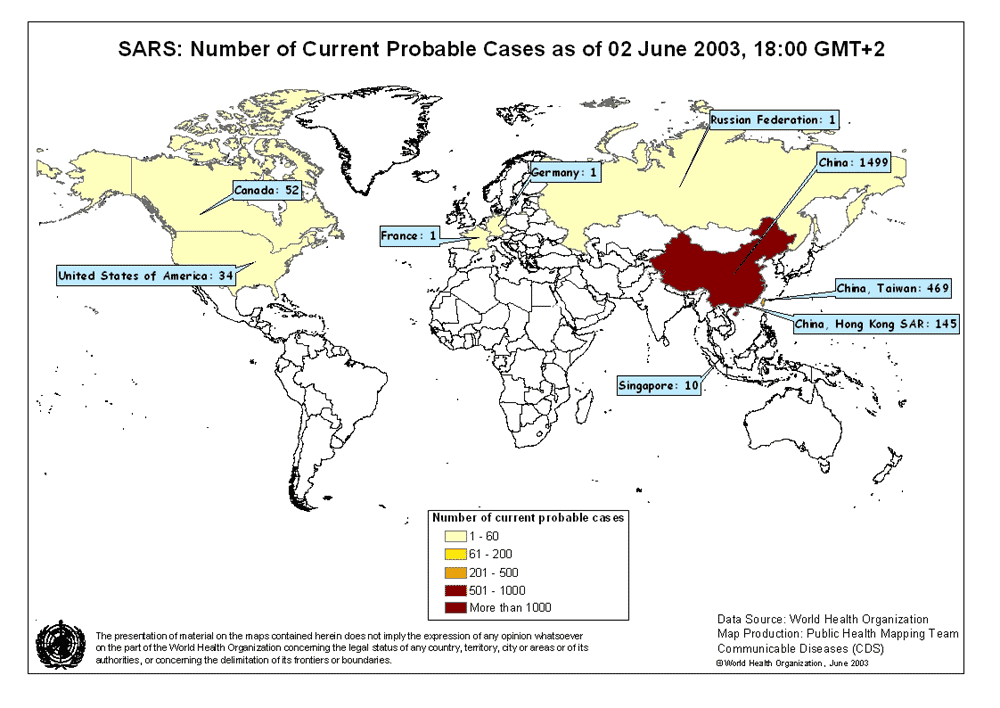 Şekil 6A
Şekil 6A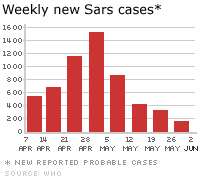 Şekil 6B
Şekil 6B