![]()

Dr Richard Hunt
VIROLOGÍA – CAPÍTULO SIETE
PARTE DIEZ
EL VIRUS DE LA INMUNODEFICIENCIA HUMANA Y EL SIDATraducido por :
Sarah M. Castillo - Jorge, Clinica Corominas
Santiago, Rep. Dominicana
VIDEOCONFERENCIA
EN INGLÉS
![]()
![]()
ENLACES A OTRAS SECCIONES DE VIH Y SIDA AL FINAL DE ESTAPAGINA
HAGA CLICK EN LOS RECUADROS GRISES PARA VENTANAS RAPIDAS CON MÁS INFORMACIÓN
HAGA CLICK EN LOS RECUADROS AMARILLOS PARA SER REDIRECCIONADO A OTROS CAPÍTULOS

La yemación causa lisis celular
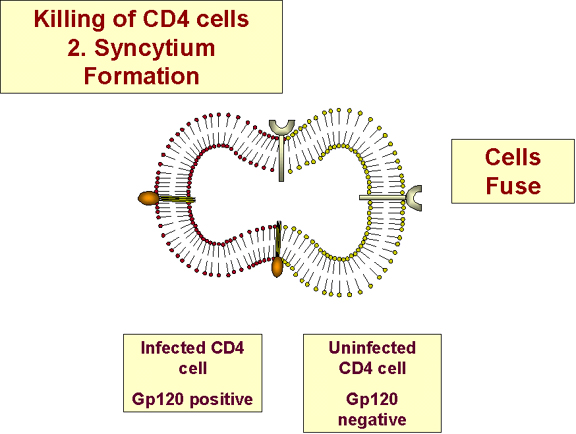
Formación de sincitios
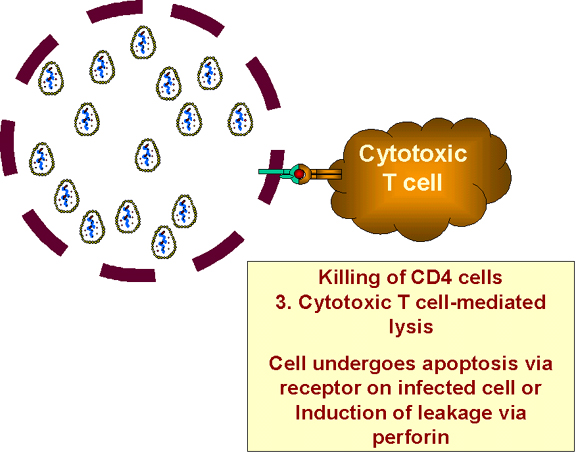
Células infectadas destruidas por células T citotóxicas
¿POR QUÉ HAY UNA PÉRDIDA PROGRESIVA DE CÉLULAS T COLABOLADORAS CD4+?
¿POR QUÉ DESAPARECEN CÉLULAS T ASESINAS CD8+ EN LAS FASES TARDIAS DE LA ENFERMEDAD?
¿Por qué, cuando tan solo 1 de 10,000 (temprano) o 1 de 40 (tardío) células muestran infección productiva, desaparecen todas las células T4? No está claro porqué todas las células CD4+ desaparecen pero hay varias posibilidades:
a) En una célula CD4 infectada, activada, gran número de viriones son sintetizados. Estos yeman de la célula y provocan rupturas membranales (figura 27). Pero la célula ha de estar infectada para que esto suceda y la mayoría de las células CD4 no son infectadas.
b) Dado que la membrana del VIH se funde con la membrana de la célula a ser infectada por un mecanismo pH-dependiente, la formación de sincitios puede ocurrir conllevando a diseminación del virus a células no infectadas (figura 27). Pero los sincitios no son muy comunes.
c) Las células infectadas que producen proteínas virales (pero no aquellas en una fase latente) presentarán esas proteínas en la superficie en asociación con antígenos del CMH clase I. La célula infectada, al igual que otras células infectadas por un virus, será destruida por las células T citotóxicas (figura 27). Pero esto sólo pasa en células que están infectadas con el VIH.
d) La Gp120 se une a la Gp41 en la superficie viral mediante interacciones no covalentes y con frecuencia es descamada de las células infectadas o de las partículas víricas. Se une a células no infectadas por el antígeno CD4. Como resultado, parecen que están infectadas y son destruidas por el sistema inmune.
e) Ha habido reportes de anticuerpos citotóxicos relacionados al SIDA en pacientes infectados que pueden reaccionar con antígenos específicos de la superficie de células T4 activadas pero no infectadas.

La unión de las citocinas induce la expresión del TNF alfa en macrófagos y la expresión del receptor en células T CD8+
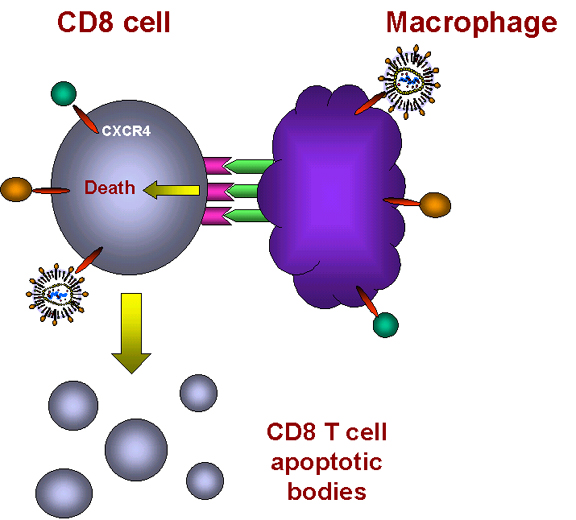
Las células se contactan entre sí y el TNF-alfa y el receptor interactúan.
Sobreviene la apoptosis
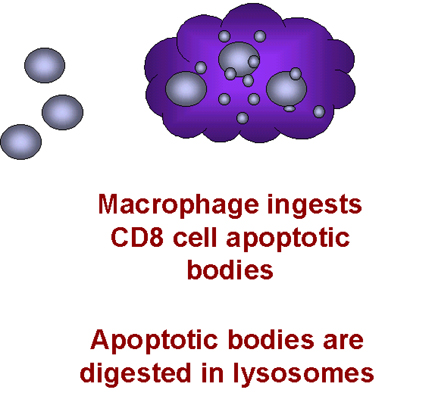
Los macrófagos integran a las células T
f) El SIDA puede tener un componente auto-inmune. En una respuesta antigénica normal llevada a cabo por las células T4, el antígeno CD4 interactúa con antígenos del CMH de clase II. Puesto que la Gp120 también se une al CD4, ésta puede simular a los antígenos del CMH de clase II ya que los dos tienen sitios de unión para el CD4; de hecho, parece que hay regiones de secuencias similares en las dos proteínas. Por tanto los anticuerpos anti-Gp120 pudieran resultar ser también anticuerpos anti-CMH. (Esto podría resultar problemático a la hora de producir una vacuna).
g) Es posible que el VIH infecte un sub-grupo de células T4 que sea vital para la propagación de toda la población de células T4
h) Las proteínas del VIH pueden alterar la función de las células T4. Hay cierta evidencia de esto.
i) Actualmente, la posibilidad más estudiada para la pérdida completa de la población de células CD4+ y CD8+ es que el VIH inicia la apoptosis en estas células (Dicha apoptosis es un proceso normal en las células T4 para superar la auto inmunidad y terminar la respuesta inmune) (figura 28). Ahora se cree que este es uno de los factores principales de pérdida de células CD4 durante la evolución de la enfermedad
Algo de lo antes mencionado puede explicar porqué sólo una minoría de las células T4 parecen infectarse en un momento determinado aún cuando todas desaparezcan en las fases tardías de la enfermedad. También podría ser que el virus intercambie la población de células T4 a medida que intercambia su co-receptor (vea arriba).
Las células CD8+ no son infectadas por el VIH (porque no poseen el receptor CD4) y el conteo de éstas permanece alto durante el curso de la enfermedad por muchos años. Y luego, algo inexplicable hasta hace poco, éstas moría rápidamente. Parece que algunos de los subtipos del VIH que se dan en fases tardías de la infección promueven una apoptosis masiva de células CD8. Aunque las células CD8 son CD4-, sí tienen el co-receptor CXCR4 y el VIH se puede unir a este (sólo las cepas inductoras de sincitio que aparecen tardíamente hacen esto). Dado que no hay antígeno CD4 presente no hay infección pero la unión al CXCR4 manda señales a la célula, la señal para la apoptosis y sobreviene un suicidio en masa de células CD8+. Es de notar, que las células CD8 sólo mueren cuando hay macrófagos presentes.¿Cómo sucede esto? Se conoce que la unión de las cepas del VIH que se encuentran en fases tardías de la infección al receptor CXCR4 activa la cascada inductora de muerte del factor de necrosis tumoral alfa (figura 28). En macrófagos, la unión del ligando al receptor CXCR4 en la superficie celular induce la expresión del TNF-alfa. En células T CD8+, esta misma unión dispara la expresión del receptor II del TNF-alfa.
Cuando tal macrófago y tal célula T CD8+ interactúan, el TNF-alfa del macrófago se une al receptor en la célula T CD8+. Esto dispara una señal de apoptosis en la célula T CD8+ resultando en la vacuolación de la célula T CD8+ (figura 28). Entonces los macrófagos fagocitan los restos celulares. Esto explica porqué los macrófagos tienen que estar presentes para que las células CD8+ mueran. ¿Por qué sucedería esto naturalmente? Porqué las quimiocinas actúan como señales de muerte para las células T CD8+? Estas células son células asesinas y pueden causar serios problemas si se dirigen al sitio equivocado. Se cree que las quimiocinas dirigen a las células T CD8+ a enfrentar una muerte mediada por macrófagos a menos que alcancen una localización apropiada.
![]()
OTRAS SECCIONES SOBRE EL VIH
PARTE I VIRUS DE LA INMUNODEFICIENCIA HUMANA Y EL SIDA
PARTE II VIH Y SIDA, LA ENFERMEDAD
PARTE III CURSO CLÍNICO DE LA ENFERMEDAD
PARTE IV PROGRESIÓN Y COFACTORES
PARTE VI SUBTIPOS Y CO-RECEPTORES
PARTE VII COMPONENTES Y CICLO DE VIDA DEL VIH
PARTE X PERDIDA DE CÉLULAS CD4
PARTE XI OTRAS CÉLULAS INFECTADAS POR EL VIH Y POLIMORFISMOS POBLACIONALES
APÉNDICE II ¿ES EL VIH CAUSA DE SIDA?
APÉNDICE III QUIMIOTERAPIA ANTI-VIH
![]()