|
x |
x |
|
 |
 |
|
x |
BAKTERIOLOGJIA |
IMUNOLOGJIA |
MIKOLOGJIA |
PARAZITOLOGJIA |
VIROLOGJIA |
|
ANGLISHT |
IMUNOLOGJIA – KAPITULLI 2
SISTEMI I
KOMPLEMENTIT
Dr Gene Mayer
Department of Pathology, Microbiology and Immunology
University of South Carolina
Columbia
Përkthyer
nga:
Dr. Burim Neziri
alergo-imunolog klinik
Asistent në
Institutin e Fiziologjisë Patologjike - Fakultetiti i Mjekësisë, Prishtinë
|
|
SPANJISHT |
|
E-MAIL DR LUL RAKA |
|
Universiteti i Prishtinës |
|
Na tregoni vlerësimin Tuaj
FEEDBACK |
|

 |
|
|
|
OBJEKTIVAT
Kuptimi i
ekzistimit të rrugëve të ndryshme të aktivizimit të SK
Njohja e
mekanizmave enzimatik dhe jo-enzimatik të aktivizimit të SK
Njohja e
karakteristikave biologjike të produkteve të aktivizimit të SK
Njohja e rëndësisë
së SK në mbrojtjen e organizmit, zhvillimin e procesit inflamator dhe
dëmtimin indor
Kuptimi i
mekanizmave rregullues të aktivizimit të komplementit dhe produktet e
tyre
 Jules Bordet
(1870-1961), zbuluesi i sistemit të komplementit.
Jules Bordet
(1870-1961), zbuluesi i sistemit të komplementit.
National Library of Medicine
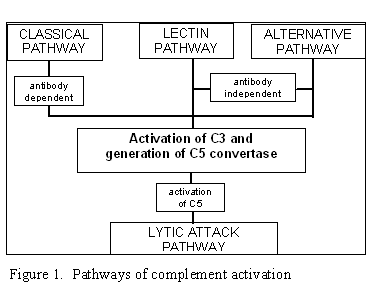
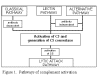 Figura 1. Rrugët e
aktivizimit të sistemit të komplementit
Figura 1. Rrugët e
aktivizimit të sistemit të komplementit |
FUNKSIONET E KOMPLEMENTIT
Historikisht, termi komplement (C) apo sistemi I komplementit (SK) është
shfrytëzuar për t’iu referuar komponentës termolabile të serumit, e cila ka qenë
e aftë që të shkatërroj bakteret (kjo komponentë e humb aktivitetitn nëse serumi
nxehet për 30 minuta në 56˚C). Megjithatë, sot dihet se sistemi i komplementit
kontribuon në mbrojtjen e organizmit edhe përmes rrugëve tjera. SK mund të
ndihmojë procesin e fagocitozës përmes
opsonizimit
të baktereve; rekrutimit dhe aktivizimit të qelizave të ndryshme, përfshirë këtu
edhe qelizat polimorfonukleare dhe makrofagët; pjesëmarrjes në rregullimin e
përgjigjes imunitare përmes antitrupave si dhe mund të ndihmoj në eliminimin e
produkteve të fundit siç janë komplekset imune dhe qelizat
apoptotike.
SK, poashtu mund të ketë efekte të dëmshme për makroorganizmin (strehuesin);
kontribuon në procesin inflamator dhe dëmtimin indor si dhe mund të provokojë
proceset anafilaktike.
SK, përbëhet nga 20
proteina të ndryshme të serumit (tab.1), të cilat prodhohen nga qeliza të
ndryshme, përfshirë hepatocitet, makrofagët dhe qelizat epiteliale të zorrëve.
Disa nga proteinat e SK lidhen për imunoglobulinat apo komponentet e membranës
qelizore. Të tjerat janë
proenzime,
të cilat, kur aktivizohen, lidhen për një apo më shumë proteina tjera të SK.
Nëse ndodhë kjo ndërlidhje, nga disa proteina të SK ndahen pjesëza (fragmente),
të cilat aktivizojnë qelizat e tjera, rrisin lëshueshmërinë vaskulare apo
shkaktojnë opsonizmin e baktereve.
|
Tabla 1. Proteinet dhe
Sistemi i Komplementit |
|
Rruga Klasike |
Rruga e Lecitinës |
Rruga Alternative |
Rruga e Lizës (Litike) |
|
Aktivizimi i proteineve:
C1qrs, C2, C3, C4
Proteinet kontrolluese:
C1-INH, C4-BP
|
Proteina mannan lidhëse (MBP), serinë –proteaza e lidhur për mannan (MASP,
MASP2) |
C3, Faktorët B & D*,
Properdina (P)
Faktorët I* & H, faktori i vonuar i akcelerimit (decay accelerating
factor - DAF), Receptori i komplementit (Complement receptor 1 -CR1),
etj.
|
C5, C6, C7, C8, C9
Proteina S
|
|
Për komponentet e nënvizuara është i
domosdoshëm ndërmjetësimi enzimatik për t’u aktivizuar.
Komponentet e shënuara me yll kanë
aktivitet nativ enzimatik. |
|
| |
RRUGËT E
AKTIVIZIMIT TË SISTEMIT TË KOMPLEMENTIT
Aktivizimi i SK mund të zhvillohet nëpërmjet katër rrugëve (Figura 1): rrugës
klasike, rrugës së lecitinës, rrugës alternative dhe rrugës së lizës apo rrugës
së atakut membranor. Rruga klasike dhe ajo alternative përcillen me aktivizimin
e C5 konvertazës dhe rezultojnë me prodhimin e komponentës C5b, e cila është
esenciale për aktivizimin e rrugës së atakut membranor.
|
|
FILM
Aktivizimi i komplementit dhe funksionet biologjike
High Resolution Quicktime
Loë Resolution Quicktime
© Scott R. Barnum, University of Alabama, Birmingham, Ala., USA and
The MicrobeLibrary
|
|
CGAP
Rrugët e detajuara të sistemit të komplementit nga CGAP/Biocarta |
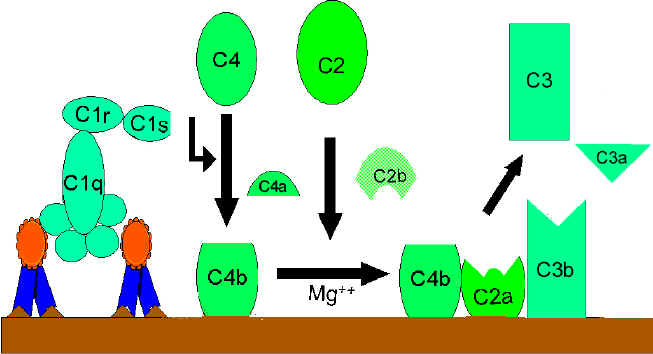
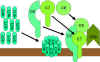
RRUGA KLASIKE (Figura 2)
Aktivizimi i
komponentës C1
C1, është
proteinë me shumë njësi e cila në vete përmban tri proteina të tjera (C1q,
C1r, C1s). Kjo proteinë lidhet për regjionin Fc të molekulës së IgG dhe IgM,
i cili regjion më parë ka hyrë në interaksion me antigjenin. Jonet e Ca++
dhe Mg++ janë të domosdoshme për realizimin e lidhjës së komponentës C1 me
antitrupat, ndërsa kjo lidhje nuk mund të ndodhë me antitrupat të cilët nuk
janë të lidhur me antigjenet. Në disa raste komponenta C1 mund të lidhet me
imunoglobulinat e grumbulluara edhe pa praninë e këtyre joneve (p.sh. IgG e
grumbulluara). Kjo lidhje mund të realizohet përmes komponentës C1q e cila
duhet gjithsesi të jetë e lidhur me së paku dy molekula të antitrupave për
të qenë fikse. Lidhja e komponentës C1q rezulton me aktivizimin e
komponentës C1r e cila më pas aktivizon komponentën C1s. Rezultati i gjithë
kësaj është formimi i komponentës C1qrs aktive, e cila në fakt paraqet një
enzim, i cili do të ndanë komponentën C4 në dy fragmente C4a dhe C4b.
Aktivizimi i
komponentës C4 dhe C2 (gjenerata e C3 konvertazës)
Fragmenti C4b i
krijuar do të lidhet për membranën qelizore derisa fragmenti tjetër C4a do
të lirohet në ambientin mikroqelizor. Komponenta ‘’C1qrs’’ do të veprojë
edhe në komponentën C2 duke e ndarë atë në dy fragmente të reja, C2a dhe
C2b. Fragmenti C2a lidhet për membranën qelizore, së bashku me fragmentin
C4b, ndërsa fragmenti C2b lirohet në ambientin mikroqelizor. Rezultat i
bashkëveprimit të fragmenteve C4bC2a është krijimi i C3 konvertazës, e cila
më pas do të vepron në komponentën C3 duke e ndarë atë në fragmentet C3a dhe
C3b.
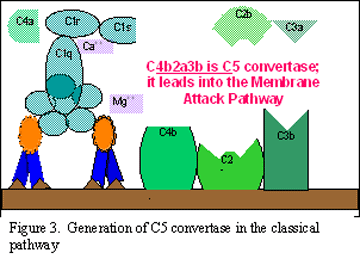
Aktivizimi i
komponetës C3 (gjenerata e C5 konvertazës)
Fragmenti C3b
së bashku me C4b dhe C2a do të lidhen për membranën qelizore, ndërsa
fragmenti C3a lirohet në ambientin mikroqelizor. Bashkëveprimi i fragmenteve
C4bC2aC3b rezulton me krijimin e C5 konvertazës e cila njëherit paraqet edhe
fundin e rrugës klasike të aktivizimit të komplementit.
Shumë produkte
të rrugës klasike tregojnë aktivitet të fuqishëm biologjik dhe në këtë
mënyrë ndihmojnë në mbrojtjen e organizmit. Disa nga këto produkte mund të
shprehin edhe efekte të dëmshme, nëse prodhimi i tyre ndodhë në mënyrë të
pakontrolluar. Në tabelën 2 janë përgjithësuar aktivitetet biologjike të
komponenteve të rrugës klasike.
|
Tabla 2. Aktiviteti biologjik i produkteve të rrugës klasike |
|
Komponenta |
Aktiviteti biologjik |
| C2b |
Prokininë;
zbërthehet nga plazmina për të krijuar kininën, e cila mund të qojë
te zhvillimi i edemës |
| C3a |
Anafilatoksina;
mund të aktivizon dhe degranulon bazofilet dhe mastocitet duke
rezultuar kështu në rritjen e permeabilitetit vaskular dhe
kontraksionin e muskujve të lëmuar, të cilat mund të qojnë deri në
gjendjen e anafilaksisë |
| C3b |
Opsoninë;
stimulon fagocitozën duke u lidhur për receptorët e sistemit të
komplementit
Aktivizon qelizat fagocituese |
| C4a |
Anafilatoksinë
(efekt të ngjashëm me C3a, por dukshëm më të dobësuar) |
| C4b |
Opsoninë;
stimulon fagocitozën duke u lidhur për receptorët e sistemit të
komplementit |
Nëse rruga klasike nuk kontrollohet më
tutje, do të vazhdojë prodhimi i fragmenteve të tjera të sistemit të
komplementit (C2b, C3a dhe C4a). Prandaj, duhet të ekzistojë patjetër një mënyrë
e cila e rregullon aktivitetin e rrugës klasike të aktivizimit të komplementit.
Tabela 3 përshkruan rrugët përmes të cilave rregullohet aktiviteti i rrugës
klasike.
|
Tabla 3. Rregullimi i Rrugës Klasike |
|
Komponenta |
Rregullimi |
|
Të gjitha |
C1-INH; ndanë (disocion) C1r dhe C1s nga C1q |
| C3a |
C3a deaktivatori (C3a-INA;karboksipeptidaza B); deaktivizon C3a |
| C3b |
Faktorët H dhe I; Faktori H nxit zbërthimin e C3b përmesë Faktorit I |
| C4a |
C3-INA |
| C4b |
C4 proteina lidhëse (C4-BP) dhe Faktor I; C4-BP nxit zbërthimin e
C4b përmesë Faktorit I; C4-BP gjithashtu parandalonë lidhjen e C2a
me C4b duke bllokuar kështu formimin e C3 konvertazës |
Rëndësia e
komponentës C1-INH në rregullimin e aktivitetit të rrugës klasike më së miri
shihet te rastet kur mungon kjo komponentë. Është vëretjur se mungesa (deficienca)
e komponetës C1 INH përcillet me zhvillimin e sëmundjes së angioedemës
hereditare.
|

A.Gjenerimi i C3 konvertazës në rrugën klasike

B. Gjenerimi i C5 konvertazës në rrugën klasike

C. Aktivizimi i C3 me rrugën klasike
Figura 2
|
 Figura 3. Rruga
e lecitinës
Figura 3. Rruga
e lecitinës |
RRUGA E
LECITINËS
Rruga e
lecitinës (Figura 3) është shumë e ngjashme me rrugën klasike. Kjo rrugë
fillon pas lidhjes së manozë-lecitinës (mannose binding lecitin-MBL) për
sipërfaqen e murit bakterial. Kjo lidhje realizohet përmes manozës e cila
përmban polisaharide dhe në momentin kur ndodh kjo lidhje do të krijohen dy
serinë proteaza, MASP1 dhe MASP2 (MBL-associated serine proteases). MASP1
dhe MASP2 luajnë rol të ngjashëm sikurse fragmentet C1r dhe C1s të rrugës
klasike, gjegjësisht MBL luanë rolin e fragmentit C1q të rrugës klasike.
Krijimi i kompleksit tremolekular MBL/MASP-1/MASP2, rezulton me aktivizimin
e MASPs dhe më tutje ndarjen e kompnentës C4 në fragmented C4a dhe C4b.
Fragmenti C4b lidhet për membranën qelizore, ndërsa fragmenti C4a do të
lirohet në ambientin mikroqelizor. MASP e aktivizuar do të veprojë
gjithashtu në komponentën C2 duke e ndarë atë në dy fragmente tjera - C2a
dhe C2b. Në vazhdim fragmenti C2a së bashku me C4b lidhen për membranën
qelizore, ndërsa C2b lirohet në ambientin mikroqelizor. Rezultat i veprimi
të kompleksit C4bC2a është krijimi i C3 konvertazës. Ky kompleks vepron në
komponetën C3 duke e ndarë atë ne dy fragmente C3a dhe C3b. Fragmenti C3b së
bashku me kompleksin C4bC2a do të lidhet për membranën qelizore, ndërsa
fragmenti C3a do të lirohet në ambientin mikroqelizor.
Aktiviteti biologjik dhe proteinat rregullatore të rrugës së lecitinës janë
tërësisht të ngjashme me ato të rrugës klasike.
|
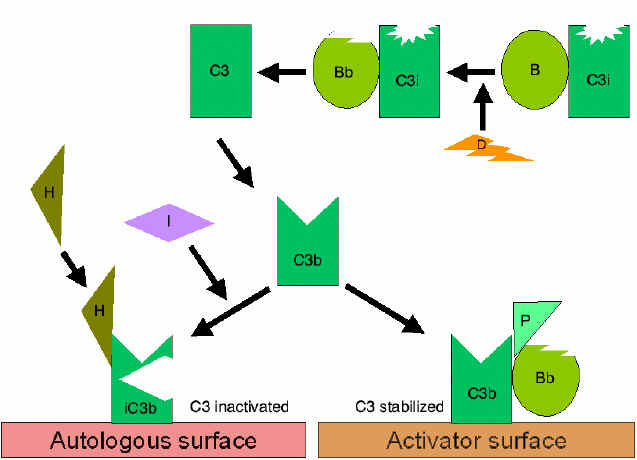
 Figura
4. Aktivizimi spontan i komponentës C3 (C3 tick-over) Figura
4. Aktivizimi spontan i komponentës C3 (C3 tick-over) |
RRUGA
ALTERNATIVE
Rruga
alternative fillon me aktivizimin e komponentës C3. Për t’u aktivizuar kjo
rrugë janë të nevojshëm faktori B, D dhe jonet e Mg++, të cilët normalisht
ndodhen në serum.
Skema
e përgjithshme e formimit të fragmentit C3b
(Figura 4)
Në kushte
normale në serum në menyrë spontane do të hidrolizohet një sasi fare e
vogël e komponetës C3 e cila rezulton me prodhimin e fragmentit C3i.
Faktori B lidhet për fragmentin C3i dhe në këtë mënyrë nxisin reaksionin
e faktorit D, i cili më pas vepron në faktorin B duke krijuar nga ai
fragmentin Bb. Kompleksi C3iBb vepron ngjashëm sikurse C3 konvertaza dhe
më pas ndan komponentën C3 në fragmentet C3a dhe C3b. Pas formimit të
fragmentit C3b, faktori B do të lidhet për të dhe ky kompleks më pas
mund të zbërthehet nga faktori D. Rezultat i veprimit të kompleksit
C3bBb është krijimi i C3 konvertazës e cila më pas do të vazhdon të
gjenerojë më shumë fragmentin C3b. Nëse ky proces vazhdon pa u
kontrolluar atëherë komponenta C3 e sistemit të komplementit do të
harxhohet në tërësi, prandaj prodhimi spontan i fragmentit C3b është i
kontrolluar në mënyrë strikte.
|

 Figura 5.
Rregullimi i aktivitetit të komponentës C3 përmes DAF
Figura 5.
Rregullimi i aktivitetit të komponentës C3 përmes DAF
 Figura
6. Rregullimi i aktivitetit të komponentës C3 përmes Cr1 Figura
6. Rregullimi i aktivitetit të komponentës C3 përmes Cr1
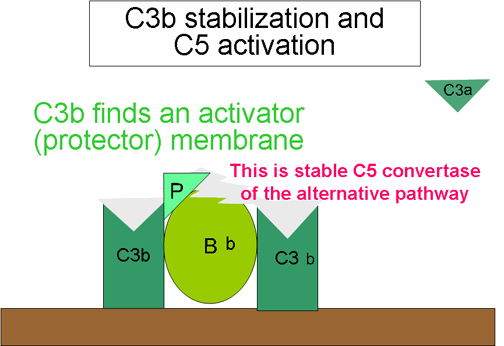
 Figura
7. Stabilizimi i C3 konvertazës Figura
7. Stabilizimi i C3 konvertazës
 Figura 8.
Stabilizimi i C5 konvertazës te rruga alternative
Figura 8.
Stabilizimi i C5 konvertazës te rruga alternative |
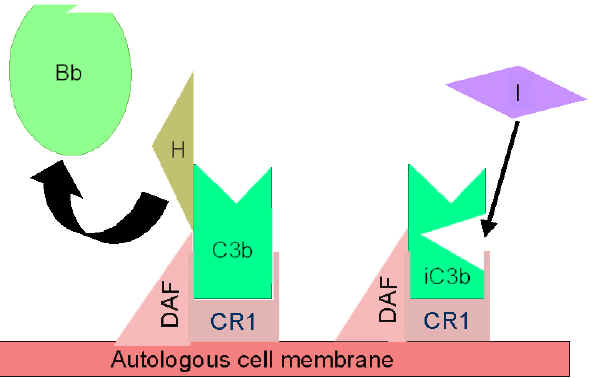
Kontrollimi i skemës së përgjithshme (Figurat 5 dhe 6)
Fragmenti
C3b i prodhuar spontanisht lidhet për membranën qelizore autologe dhe
hyn në interaksion me DAF (decay accelerating factor) dhe kështu do të
bllokohet lidhja e faktorit b me fragmentin C3b dhe rrjedhimisht edhe
formimi i sasive te reja të C3 konvertazës. Për më shumë, DAF nxit
zbërthimin e fragmentit Bb nga komponenta tjetër C3b, të cilat kanë
ndikuar në formimin e C3 konvertazës dhe në këtë mënyrë e ndalon
krijimin e mëtutjeshëm të C3b. Disa qeliza posedojnë receptorët 1 për
sistemin e komplementit (CR1). Lidhja e C3b me CR1 nxit zbërthimin
enzimatik të fragmentit C3b përmes faktorit I. Lidhja e C3 konvertazës
(C3bBb) për CR1 gjithashtu do të ndikojë në disocimin e fragmentit Bb
nga kompleksi. Prandaj mund të themi se ato qeliza të cilat e posedojnë
CR1 luajnë rol në kontrollimin e skemës së përgjithshme të krijimit të
C3 konvertazës. Së fundi, faktori H mund të lidhet për fragmentin C3b i
cili mund të jetë i lirë apo i lidhur për qelize dhe kështu të nxit
zbërthimin enzimatik të fragmentit C3b përmes faktorit I. Nga ajo që u
tha më sipër mund të konkludohet se kontrollimi i skemës së përgjithshme
të krijimit të C3 konvertazës mund të bëhet përmes bllokimit të krijimit
të C3 konvertazës, disocimit (zbërthimit) të C3 konvertazës ose përmes
zbërthimit enzimatik të fragmentit C3b. Rëndësia e kontrollimit të kësaj
skeme më së miri është ilustruar te pacientët me deficiencë gjenetike të
faktorit H apo I. Te këta pacientë, deficienca e komponentës C3
përcillet me mundësi të rritura për infeksione të ndryshme.
Stabilizimi i C konvertazës përmes aktivatorit sipërfaqësor
(Figura 7)
Në momentin
kur komponenta C3b do të lidhet për aktivatorin përgjegjës të rrugës
alternative, njëkohësisht do të lidhet edhe për faktorin B, i cili në
mënyrë enzimatike është i lidhur edhe me faktorin D dhe e gjithë kjo
lidhje rezulton me formimin e C3 konvertazës (C3bBb).
Komponenta
C3b është mjaft rezistente ndaj zbërthimit nga faktori I, prandaj C3
konvertaza nuk zbërthehet menjëherë dhe kësaj i kontribon edhe
stabilizimi nga aktivatori sipërfaqësor. Stabilizimit të kompleksit i
ndihmon edhe lidhja me C3bBb përmes properdinës. Aktivatorët e rrugës
alternative janë komponente sipërfaqësore të agjensëve patogjenë, siç
janë: LPS e bakterieve Gram negative, murit bakterial të disa baktereve
tjera dhe kërpudhave. Lidhja e C3b për aktivatorin sipërfaqësor, ndikon
që C3 konvertaza e formuar të stabilizohet dhe të vazhdoj krijimi i
sasive të reja të komponentës C3a dhe C3b, përmesë zbërthimit të
komponetës C3.
Gjenerimi i C5 konvertazës (Figura 10)
Një pjesë e
komponentës C3b e cila gjenerohet nga ndërlidhja dhe stabilizimi i C3
konvertazës me aktivatorin sipërfaqësor, lidhet me kompleksin C3bBb dhe
formon një kompleks të ri C3bBbC3b. Ky kompleks i ri paraqet C5
konvertazën e rrugës alternative. Gjenerimi (krijimi) i C5 konvertazës
paraqet edhe fundin e rrugës alternative. Rruga alternative mund të
aktivizohet nga një numër i madh i baktereve Gram-negative (më të
shpeshtat: Neisseria meningitidis and Neisseria gonorrhoea),
disa bakterie Gram-pozitive, një numër i caktuar virusësh dhe parazitësh
dhe kjo rezulton me lizën e këtyre organizmave (mikroorganizmave).
Prandaj, rruga e aktivizimit të komplementit ofron një mundësi tjetër
për mbrojtjen e organizmit nga lloje të ndryshme të mikroorganizmave
patogjen, para se të fillojë veprimi i antitrupave. Deficienca e
komponentës C3 rezulton me rritjen e ndjeshmërisë ndaj mikroorganizmave
të lartpërmendur. Rruga alternative e aktivizimit të komplementit mund
të konsiderohet si rrugë më primitive derisa rruga klasike dhe ajo e
lecitës me gjasë kanë prejardhje nga kjo rrugë.
|
| |
|
| |
Duhet të kujtojmë
se rruga alternative paraqet vijën e parë të mbrojtjes ndaj agjensëve infeksiozë
dhe njëherit mënyrën jospecifike të rezistencës dhe mbrojtës nga infeksioni duke
përjashtuar participimin e antitrupave.
Shumë baktere
Gram-negative dhe disa
Gram-pozitive, shumë virusë, parazitë dhe eritrocite
heterologe agregojnë imunoglobulinat (veçanërisht IgA) dhe disa proteina të
tjera (p.sh. proteazat, produktet e rrugës së koagulimit) dhe në këtë mënyrë
mund të aktivizojnë rrugën alternative. Proteina, cobra venom factor (CVF) është
studiuar veçanërisht në kuptimin e aktivizimit të rrugës alternative.
|
 Figura
9. Rruga e lizës (litike)
Figura
9. Rruga e lizës (litike) |
RRUGA E ATAKUT
MAMBRANOR (LIZËS) (Figura 9)
C5 konvertaza e
cila mund të ketë prejardhjen nga rruga klasike (C42a3b), e lecitinës (C42a3b)
ose ajo alternative (C3bBb3b), vepron në komponentën C5 duke e ndarë atë në C5a
dhe C5b. C5a mbetet në fazën fluide, ndërsa C5b menjëherë hyn në reaksion me C6
dhe C7 dhe ngjitet (insertohet) për membranë. Më pas, aktivizohet komponenta C8
e cila më tutje stimulon aktivizimin e disa molekulave të C9. Këto molekula
formojnë pore në membranën qelizore, përmes të cilave përmbajtja qelizore
derdhet jashtë dhe kështu shkaktohet liza qelizore. Liza qelizore nuk është
proçes enzimatik dhe mendohet se paraqitet si pasojë e dëmtimit fizik të
membranës qelizore. Kompleksi i përbërë nga C5bC6C7C8C9 quhet ndryshe edhe
kompleks i atakut membranor (membrane attack complex-MAC).
Fragmenti C5a, i
cili krijohet përmes rrugës së lecitinës shpreh disa efekte të fuqishme
biologjike. Ky fragment është një ndër
anafilatoksinet
më të fuqishme.
Gjithashtu, karakterizohet me efekte të kemotaksisë për neutrofilet dhe stimulon
proçeset respiratore në këto qeliza si dhe stimulon makrofagët për prodhimin e
citokineve. Aktiviteti i këtij fragmenti kontrollohet nga inaktivizimi i
karboksipeptidazës B (C3-INA).
Një pjesë ë
kompleksit të formuar C5b67, mund të ndahet nga membrane qelizore dhe të futet
në fazën flutide, të lidhet për qelizat qe ndodhen atypari dhe të shkaktojë
lizën e tyre. Dëmtimi i këtyre qelizave normalisht parandalohet nga Proteina S
(vitronectin), e cila lidhet për kompleksin e tillë të lirë C5b67 dhe e okupon
atë duke pamundësuar kështu lidhjen për qeliza tjera.
|
 Figura
10. Rregullimi i komponentës C1rs (C4 konvertazës) përmes C1-INH Figura
10. Rregullimi i komponentës C1rs (C4 konvertazës) përmes C1-INH
|
PRODUKTI AKTIV
BIOLOGJIK I AKTIVIZIMIT TË KOMPLEMENTIT
Aktivizimi i
komplementit rezulton me prodhimin e shumë molekulave aktive biologjike të cilat
mund të kontribojnë në rezistencën e organizmit,
anafilaksinë dhe proçesin
inflamator.
Prodhimi i
kininës
Fragmenti C2b i
cili krijohet nga rruga klasike e aktivizimit të komplementit, është një
prokininë e cila mund të kalon në formën e saj aktive biologjike duke iu
nënshtruar transformimit enzimatik nga plazmina. Teprica eventuale e C2b mund të
inaktivizohet përmes C1 inhibitorit (C1-INH), i cili quhet edhe serpinë.
Normalisht ky faktor bën zhvendosjen e C1rs nga C1qrs kompleksi (Fig.10).
Deficienca gjenetike e C1-INH rezulton me mbiprodhimin e fragmentit C2b dhe kjo
shoqërohet me paraqitjen e edemës angioneurotike hereditare. Kjo gjendje mund të
trajtohet me Danazol, i cili stimulon prodhimin e C1-INH apo me ε-acidin amino
kaproik i cili zvoglon aktivitetin e palzminës.
|
|
BURIMET E INTERNETIT
Angioedema trashëguese
Trashëgimia Mendeliane on-line (NIH)
|
Anafilatoksinat
C4a, C3a dhe C5a (në
gjendje të hiperaktivitetit) të gjitha janë anafilatoksine të cilat shkaktojnë
degranulimin e bazofileve dhe mastociteve si dhe kontraksionin e muskujve të
lëmuar. Efektet e padëshiruara të këtyre peptideve i parandalon dhe kontrollon
karboksipeptidaza B ( C3a-INA).
Faktorët
kemotaktik
C5a dhe MAC (C5b67)
janë faktorë kemotaktik. C5a poashtu është një faktor i fuqishëm i aktivizimit
të neutrofileve, bazofileve dhe makrofagëve e gjithashtu mund të indukojë edhe
adhezionin (ngjitjen) e molekulave për qelizat endoteliale vaskulare.
Opsoninet
C3b dhe C4b të
ngjitura për sipërfaqen membranore të mikroorganizmave, lidhen për CR1 e
qelizave fagocituese dhe kështu provokojnë fagocitozën e tyre.
Produktete tjera
aktive biologjike të aktivizimit të sistemit të komplementit
Prodhimet e
zbërthimit të komponentës C3 (iC3b, C3d dhe C3e) përmes receptorëve të caktuar
gjithashtu mund të lidhen për qeliza të ndryshme dhe kështu të modulojnë
funksionin e tyre.
Në përgjithësi mund
të themi se sistemi i komplementit merr pjesë gjithashtu në mbrojtjen specifike
dhe jospecifike të organizmit dhe gjeneron (krijon) shumë produkte biologjike të
cilat mund të kenë edhe rol të rëndësishëm patofiziologjik (tabela 4).
Deri sot njihen
shumë gjendje të deficiencave gjenetike kryesisht të komponentës C të sistemit
të komplementit, por deficiencat e komponetës C3 janë më seriozet dhe ndonjëherë
edhe fatale. Deficiencat e sistemit të komplementit gjithashtu mund të vërehen
te sëmundjet imunokomplekse ( p.sh. SLE) si dhe te infeksionet bakterore akute
dhe kronike, infeksionet virale dhe ato me parazitë.
|
|
|
|
|
|
|
Tabla 4. Aktiviteteet e prodhimeve të aktivizuara të SK dhe
kontrolli I tyre |
|
Fragmenti |
Aktiviteti |
Efekti |
Faktori kontrollues |
| C2a |
Prokininë, grumbullimin e lëngut |
Edemën |
C1-INH |
| C3a |
Degranulimin e bazofileve dhe mastociteve; rrit permeabilitetit
vaskular dhe kontraksionin e muskujve të lëmuar |
Anafilaksinë |
C3a-INA |
| C3b |
Opsoninë, aktivizimi i qelizave fagocituese |
Fagocitozën |
Faktorët H dhe I |
| C4a |
Degranulimin e bazofileve dhe mastociteve; rrit permeabilitetin
vaskular dhe kontraksionin e muskujve të lëmuar |
Anafilaksinë
(formë
më të lehtë) |
C3a-INA |
| C4b |
Opsoninë |
Fagocitozën |
C4-BP dhe Faktori I |
| C5a |
Degranulimin e bazofileve dhe mastociteve; rrit permeabilitetin
vaskular dhe kontraksionin e muskujve të lëmuar |
Anafilaksinë
(formë
të rëndë) |
C3a-INA |
|
Kemotaksën, stimulon proceset respiratore, aktivizon qelizat
fagocituese, stimulon prodhimin e citokineve |
Inflamacionin |
| C5bC6C7 |
Kemotaksën |
Inflamacionin |
Protein S (vitronektina) |
|
Ngjitja për membranën qelizave tjera |
Dëmtimi indor |
|
|
Ju keni mësuar:
Proteinat e
sistemit të komplementit
Dallimet dhe
ngjashmëritë në rrugët e ndryshme të aktivizimit të C3
Rolin e rrugëve të
ndryshme të aktivizimit në imunitetin specifik dhe jospecifik
Rolin e prodhimeve të ndryshme të aktivizimit të SK në fuqizimin e
imunitetit specifik dhe jospecifik dhe në inflamacion |
|
Tabla 5. Sëmundjet
dhe deficiencat e komplementit |
|
Rruga/Komponenta |
Sëmundja |
Mekanizmi |
|
Rruga Klasike |
|
| C1INH |
Angioedema Hereditare |
Mbiprodhimi i C2b (prokinina) |
| C1, C2, C4 |
Predispozicion për SLE |
Opsonizimi i komplekseve imune ndikonë që ato të jenë më të
tretshme dhe deficinenca ndikonë në rritjen e percipitimit në
inde dhe provokimin e inflamacionit |
|
Rruga e Lectinës |
|
| MBL |
Ndjeshmëri për infeksione të shpeshta te të porsalindurit dhe te
përsonat imunosupresiv |
Paaftësia për të nxitur (filluar) rrugën e lecitinës |
|
Rruga Alternative |
|
|
Faktorët B apo D |
Ndjeshmëri për infeksione bakteriale piogjene |
Mangësia në procesin e opsonizimit të bakterieve |
| C3 |
Ndjeshmëri për infeksione bakteriale |
Mangësia në procesin e opsonizimit dhe paftësi për të
konkretizuar atakun membranorë |
| C5, C6, C7 C8, and C9 |
Ndjeshmëri për infeksione me bakterie gram-negative |
Paftësia për të sulmuar membranën e jashtme të bakterieve
gram-negative |
|
Properdina (E lidhur për X) |
Ndjeshmëri e rritur për meningjitin menigokocigela |
Mangësia në procesin e opsonizimit të bakterieve |
| Faktorët
H apo I |
C3 deficiencë dhe ndjeshmëri për infeksione bakteriale |
Aktivizim i pakontrolluar i kompnentës C3 përmesë rrugës
alternative e cila rezulotnë me harxhim të kompnentës C3 |
|
|
|
 Kthehu
në seksionin e imunologjia të Microbiology and Immunology On-line
Kthehu
në seksionin e imunologjia të Microbiology and Immunology On-line
faqja së fundi e ndërruar në
Sunday, February 15, 2015
faqja mbahet nga
Richard Hunt
|
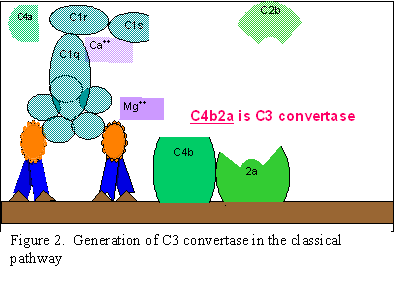
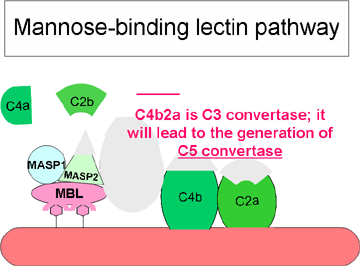 Figura 3. Rruga
e lecitinës
Figura 3. Rruga
e lecitinës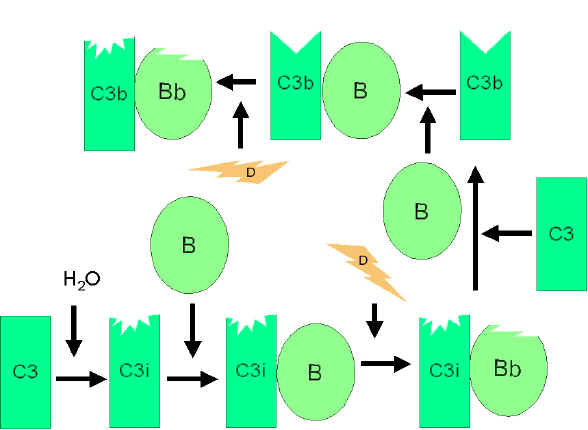 Figura
4. Aktivizimi spontan i komponentës C3 (C3 tick-over)
Figura
4. Aktivizimi spontan i komponentës C3 (C3 tick-over)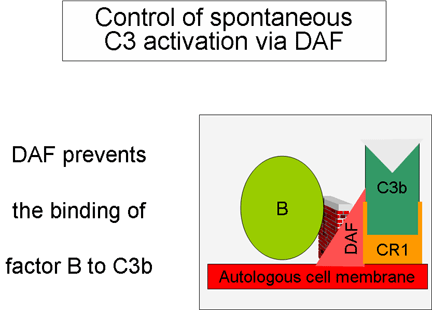
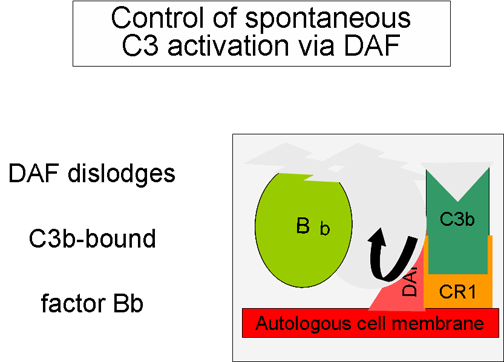 Figura 5.
Rregullimi i aktivitetit të komponentës C3 përmes DAF
Figura 5.
Rregullimi i aktivitetit të komponentës C3 përmes DAF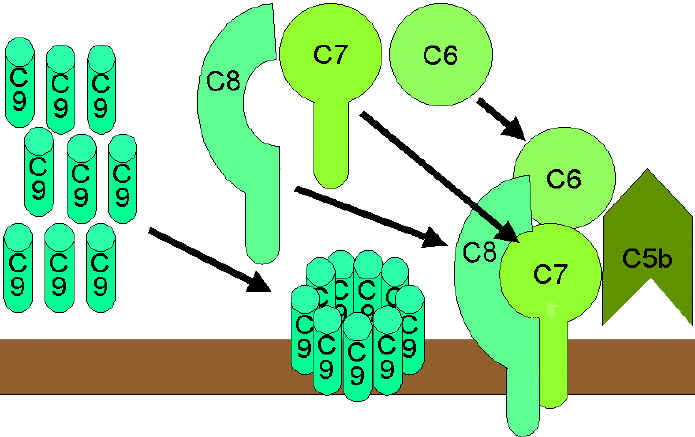 Figura
9. Rruga e lizës (litike)
Figura
9. Rruga e lizës (litike)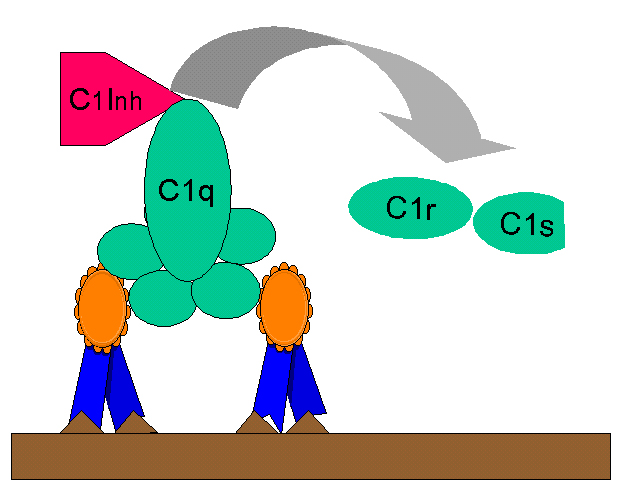 Figura
10. Rregullimi i komponentës C1rs (C4 konvertazës) përmes C1-INH
Figura
10. Rregullimi i komponentës C1rs (C4 konvertazës) përmes C1-INH