| x | x | ||||
|
|
 |
||||
| BACTERIOLOGIE | IMMUNOLOGIE | MYCOLOGY | PARASITOLOGY | VIROLOGIE | |
| VIDEO LECTURE |
|
||||
| EN ANGLAIS | |||||
| EM PORTUGUES - PORTUGUESE | |||||
|
Let us know what you think FEEDBACK |
|||||
| SEARCH | |||||
|
|
|||||
| Logo image © Jeffrey Nelson, Rush University, Chicago, Illinois and The MicrobeLibrary | |||||
|
LECTURES Male et al. Immunology 7ème édition chapitre 7 et pages 40-4 |
|||||
|
OBJECTIFS DU COURS MOTS-CLÉS
|
COMPARAISON DES BCR ET DES
TCR
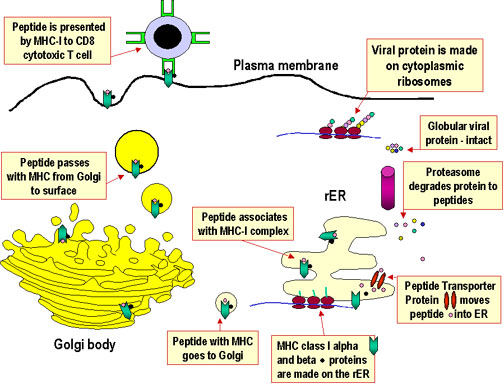
L’apprêtement de l'antigène et sa présentation sont des processus qui se produisent dans une cellule et qui conduisent à la fragmentation (protéolyse) de protéines, à l'association des fragments générés à des molécules du CMH, et à l'expression des complexes peptide-CMH à la surface des cellules où ils peuvent être reconnus par le récepteur de la cellule T. Toutefois, le cheminement menant à l'association de fragments de protéines avec des molécules du CMH différent pour les molécules du CMH de classe I et de classe II. Les molécules du CMH de classe I présentent des produits de dégradation de protéines intracellulaires (endogènes) générés dans le cytosol. Les molécules du CMH de classe II présentent des fragments dérivés de protéines extracellulaires (exogènes) qui sont générés dans un compartiment intracellulaire.
|
||||
|
|
|
||||
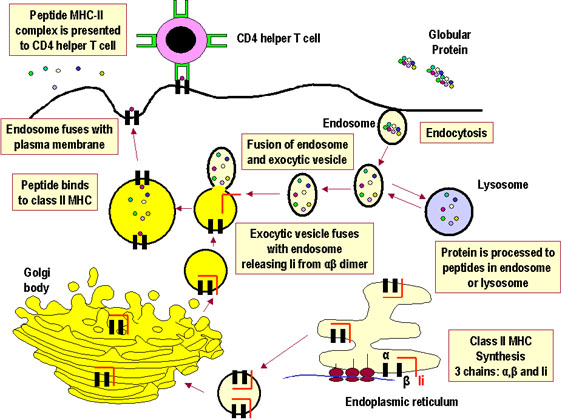 Figure 2
Figure 2Apprêtement des antigènes exogènes en vue de leur présentation par les molécules du CMH de classe II. |
|||||
|
|
|
||||
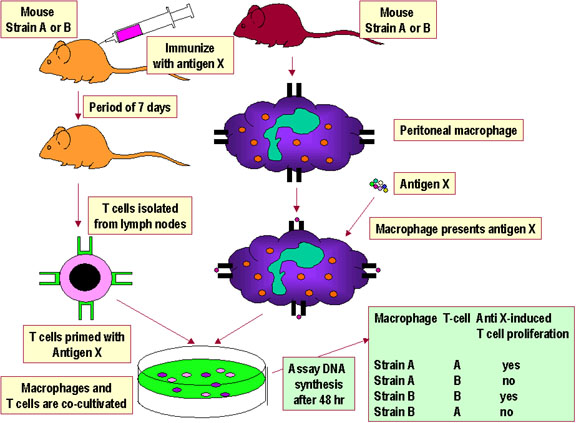 Figure 3
Figure 3Restriction au CMH des interactions Thelper/APC. |
|||||
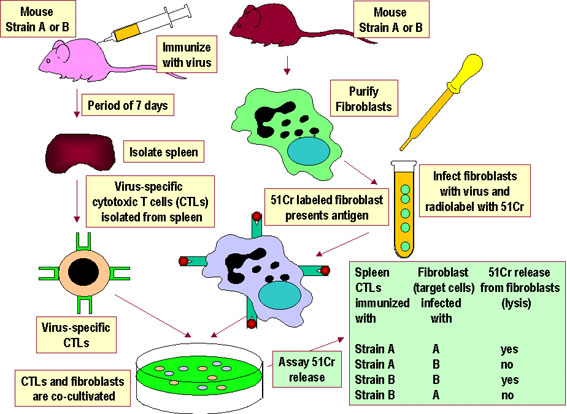 Figure 4
Figure 4Des CTL spécifiques d’un virus provenant d’une lignée A ou d’une lignée B de souris ne pourront lyser que les cellules cibles syngéniques infectées par ce virus. Ces CTL ne lyseront pas des cellules non-infectées et ne seront pas alloréactives. Plus généralement, les CTL et les cellules cibles doivent provenir du même animal ou d’animaux partageant le même allèle du CMH de classe I pour que les cellules cibles présentent l’antigène viral aux CTL. |
RESTRICTION AU CMH DU SOI Pour qu’une cellule T reconnaisse et réponde à un antigène protéique étranger, elle doit reconnaître le CMH de la cellule présentatrice comme CMH du soi. C'est ce qu'on appelle la restriction au CMH. Les lymphocytes T auxiliaires reconnaissent l'antigène dans le contexte de molécules du CMH de classe II du soi. Les lymphocytes T cytolytiques reconnaissent l'antigène dans le contexte des molécules du CMH de classe I du soi. Le processus par lequel les cellules T deviennent restreintes au molécules du CMH du soi se produit dans le thymus. Les systèmes expérimentaux ayant
permis de démontrer la restriction au CMH du soi pour les cellules T
auxiliaires et pour les cellules T cytotoxiques sont présentés dans les
Figures 3 et 4, respectivement. |
||||
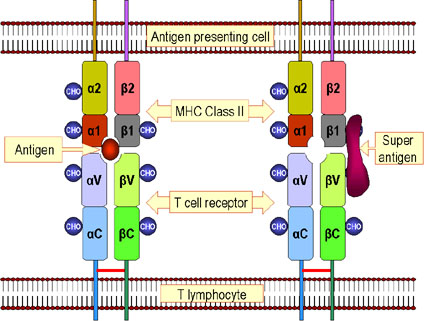
Figure 5 |
CELLULES PRÉSENTATRICES D’ANTIGÈNES Les trois principaux types de cellules présentatrices d'antigène sont les cellules dendritiques, les macrophages et les cellules B. Il existe d'autres cellules qui expriment des molécules du CMH de classe II (comme par exemple les cellules épithéliales thymiques) et qui peuvent agir en tant que cellules présentatrices d'antigène dans certains cas. Les cellules dendritiques, qui se trouvent dans les tissus comme la peau ou autres, peuvent ingérer des antigènes par pinocytose et les transporter vers les ganglions lymphatiques et la rate. Dans les ganglions lymphatiques et la rate, ces cellules se retrouvent principalement dans les zones où sont présentes les cellules T. Les cellules dendritiques sont les cellules présentatrices d'antigènes les plus efficaces et peuvent présenter les antigènes à des cellules T naïves (ou vierges). En outre, elles peuvent présenter des antigènes internalisés en association soit avec les molécules du CMH de classe I (phénomène appelé « présentation croisée ») ou du CMH de classe II, et ce, bien que la voie prédominante de présentation d’antigènes internalisés est la voie du CMH de classe II. Le second type de cellule présentant l'antigène est le macrophage. Ces cellules ingèrent l’antigène par phagocytose ou pinocytose. Les macrophages ne sont pas aussi efficaces dans la présentation de l'antigène aux lymphocytes T naïfs, mais ils sont très efficaces dans l'activation des cellules T mémoires. Le troisième type de cellule présentant l'antigène est la cellule B. Ces cellules se lient à l’antigène via leur immunoglobuline de surface et ingèrent ainsi l’antigène par pinocytose. Comme les macrophages, ces cellules ne sont pas aussi efficaces que les cellules dendritiques dans la présentation de l'antigène aux lymphocytes T naïfs. Les lymphocytes B sont par contre très efficaces pour la présentation de l'antigène aux cellules T mémoires, en particulier lorsque la concentration de l'antigène est faible, car l'immunoglobuline de surface sur les cellules B se lie à l'antigène avec une affinité élevée.
Les superantigènes sont des antigènes qui peuvent
activer des lymphocytes T (voir le chapitre « antigènes ») de façon
polyclonale pour leur permettre de produire de grandes quantités de
cytokines (ce qui peuvent avoir des effets pathologiques). Ces antigènes
doivent être présentés aux lymphocytes T en association avec des molécules
du CMH de classe II, mais, dans ce cas, l'antigène n'a pas besoin d'être
apprêté. La Figure 5 compare la façon dont les antigènes classiques et les
superantigènes sont présentés aux cellules T. Dans le cas d'un superantigène,
la protéine antigénique intacte se lie à des molécules du CMH de classe II
et à une ou plusieurs régions Vbeta du TCR. L'antigène n'est pas lié à
l’intérieur du site de liaison peptidique de la molécule de CMH ni au site
de liaison d'antigène du TCR. Ainsi, toute cellule T qui utilise une chaîne
Vbeta particulière du TCR sera activé par un superantigène donné, ce qui
entraîne l'activation d'un grand nombre de cellules T. Chaque superantigène
va se lier à un ensemble différent de régions Vbeta. |
||||
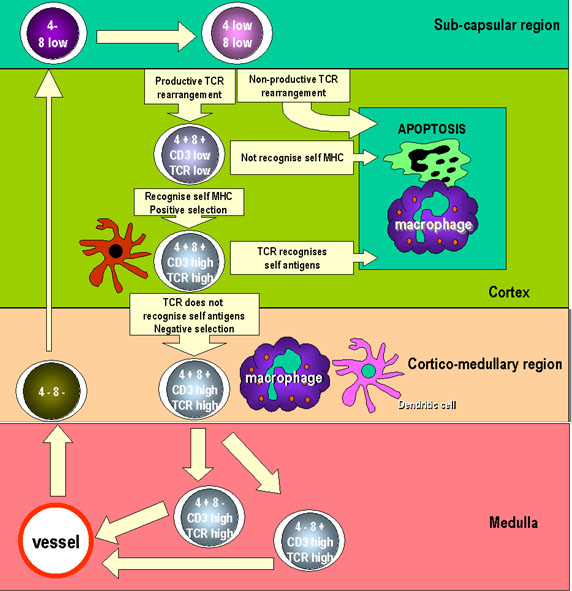 Figure 6
Figure 6Les cellules T pré-thymiques pénètrent dans le thymus et prolifèrent sous la forme de grands lymphoblastes dans la région sous-capsulaire du thymus. Les lymphoblastes se divisent pour former un pool de cellules qui se différencient. A ce stade, les cellules n’expriment les marqueurs CD4 et CD8 que très faiblement. Les gènes du TCR sont réarrangés et leurs produits sont aussi exprimés assez faiblement. Au fur et à mesure que les cellules maturent, elles migrent vers le cortex où elles interagissent avec les cellules épithéliales corticales qui forment un réseau offrant de larges surfaces d’interaction avec les autres cellules. Les TCR exprimés à la surface des thymocytes interagissent avec les molécules du CMH exprimées par les cellules épithéliales thymiques corticales, conduisant à la sélection positive. Les cellules qui ne sont pas sélectionnées sont alors soumises à une apoptose suivie d’une phagocytose par les macrophages par exemple. Dans le cortex les cellules expriment plus fortement CD3, CD4, CD8 et le TCR. Les thymocytes porteurs de TCR auto-réactifs sont éliminés par contact avec des auto-antigènes exprimés par les des cellules dendritiques et des macrophages : c’est la sélection négative. Des cellules exprimant soit CD4 soit CD8 apparaissent et migrent vers la périphérie en utilisant des vaisseaux spécialisés de la région cortico-médullaire du thymus. |
EDUCATION THYMIQUE Les deux types de cellules T,
Thelper et Tcytotoxiques sont restreintes au CMH du soi. Par ailleurs, les
cellules T ne reconnaissent normalement pas les auto-antigènes. Comment les
cellules T deviennent-elles restreintes au CMH du soi et pourquoi les
cellules T auto-réactives ne sont-elles pas produites? On s'attend à ce que
des réarrangements VDJ faits au hasard dans les cellules T puissent générer
des cellules T capables de reconnaître des molécules du CMH du non soi et
aussi à ce que certains lymphocytes T soient capables de reconnaître les
antigènes du soi. C'est le rôle du thymus de garantir que les seules
cellules T qui arrivent en périphérie sont restreintes au CMH du soi et sont
incapables de réagir avec des auto-antigènes. Les cellules T fonctionnelles
présentes en périphérie doivent reconnaître les antigènes étrangers associés
au CMH du soi : en effet, les APC ou les cellules cibles présentent
l’antigène étranger associé au CMH du soi. Cependant, on n'a pas besoin de
cellules T fonctionnelles en périphérie qui reconnaissent des antigènes (du
soi ou étrangers) associé à un CMH étranger. On n’a pas non plus besoin de
cellules T fonctionnelles en périphérie qui peuvent reconnaître les
antigènes du soi associés aux CMH du soi parce que ces cellules pourraient
infliger des dommages à nos tissus normaux sains. Initialement, la cellule pré-T qui
entre dans le thymus est CD4-CD8-. Dans le thymus, cette cellule devient CD4
+ CD8 +, puis, à la suite de la sélection positive et négative, la cellule
devient soit un T CD4 + ou un T CD8 +. L'engagement vers la lignée T CD4 +
ou T CD8 + dépend de la classe de molécule du CMH que rencontre la cellule
T. Si une cellule T CD4 + CD8 + reconnaît une molécule de classe I,
l’expression de CD4 va être perdue et la cellule va devenir une cellule T
CD8. Si une cellule T CD4 + CD8 + reconnaît une molécule de classe II,
l’expression de CD8 va être perdue et la cellule va devenir une cellule T
CD4+ (Figure 7). La sélection positive et négative
dans le thymus n'est pas un procédé efficace à 100%. En outre, tous les
antigènes du soi ne peuvent être exprimés dans le thymus. Ainsi, certaines
cellules T auto-réactives peuvent se rendre en périphérie. Il existe des
mécanismes supplémentaires qui sont conçus pour éliminer les cellules T
auto-réactives de la périphérie. Ces questions seront abordées dans le
chapitre sur la tolérance. |
||||
|
Les thymocytes précurseurs CD4-CD8- deviennent doubles positifs CD4 + CD8 + et expriment de faibles niveaux de chaînes alpha et bêta du récepteur à l’antigène des cellules T (TCR). La sélection positive par les molécules du CMH de classe I ou de classe II du soi se produit dans l'épithélium cortical. La majorité des cellules sont non-sélectionnées et meurent par apoptose. Les cellules restantes vont pouvoir soit interagir avec le CMH de classe I et perdront leur antigène CD4 soit avec le CMH de classe II et perdront leur antigène CD8. Les cellules auto-réactives sont ensuite éliminées à la suite de leur interaction avec des peptides antigéniques du soi qui sont présentés par des cellules présentes à la jonction cortico-médullaire et dans la médulla du thymus |
|||||
|
|
|||||
|
|
|
||||