|
x |
x |
|
 |
 |
|
DOENÇAS
INFECCIOSAS |
BACTERIOLOGIA |
IMUNOLOGIA |
MICOLOGIA |
PARASITOLOGIA |
VIROLOGIA |
|
|
IMUNOLOGIA – CAPÍTULO ONZE
RESPOSTA
AO ANTÍGENO: PROCESSAMENTO E APRESENTAÇÃO
RESTRIÇÃO AO MHC E PAPEL DO TIMO
Dr.
Gene Mayer
Professor Emeritus
University of South Carolina School of Medine
Tradução:
PhD. Myres Hopkins
|
|
EM INGLÊS |
|
|
Dê a
sua opinião
CONTATO |
|
BUSCA |
|
|
E-MAIL
DR MYRES HOPKINS |
|
ESCOLA DE MEDICINA DA
UNIVERSIDADE DA CAROLINA DO SUL |

 |
|
|
|
OBJETIVOS
Comparar e diferenciar antígenos reconhecidos pelo TCR e BCR.
Descrever as vias envolvidas no processamento de antígenos endógenos e
exógenos
Discutir restrição do MHC ao próprio na apresentação de antígenos às
células T
Descrever as principais células apresentadoras de antígenos.
Comparar e diferenciar a apresentação de antígenos convencionais e
superantígenos.
Discutir o papel da seleção positiva e negativa no timo na geração de
células T com restrição do MHC ao próprio.
PALAVRAS-CHAVE
Antígeno endógeno
Via do processamento de antígeno de classe I
Proteossomo
Transportador
Antígeno exógeno
Via do processamento de antígeno de classe II
Cadeia invariante
Restrição do MHC ao próprio
Seleção positiva
Seleção negativa
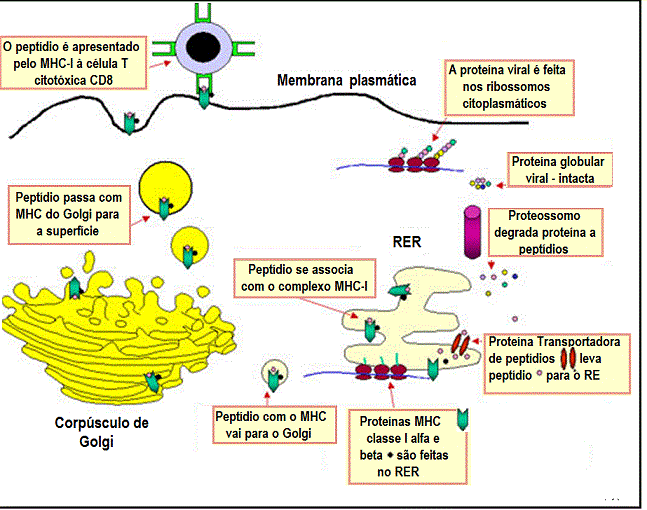
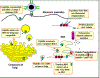 Figura 1
Figura 1
Via da apresentação de um antígeno sintetizado endogenamente com
restrição ao MHC classe I. Um exemplo de tal antígeno seria uma proteína
viral produzida na célula como resultado de uma infecção |
ComparAÇÃO ENTRE BCR E TCR
Células B e células T reconhecem diferentes substâncias como
antígenos e reconhecem de uma forma diferente. A célula B usa a
imunoglobulina ligada à superfície da célula como um receptor e
a especificidade deste receptor é a mesma da imunoglobulina que
ela é capaz de secretar após a ativação. Células B reconhecem os
seguintes antígenos na forma solúvel: 1)
Contrariamente, a esmagadora maioria dos antígenos de células T
são proteínas, e estas precisam ser fragmentadas e reconhecidas
em associação com produtos do MHC expressos na superfície de
células nucleadas, não em forma solúvel. Células T estão
agrupadas funcionalmente de acôrdo com a classe de moléculas de
MHC que se associa com os fragmentos peptídicos da proteína:
células T auxiliares reconhecem apenas aqueles peptídios
associados com moléculas de MHC classe II, e células T
citotóxicas reconhecem apenas aqueles peptídios associados com
moléculas de MHC classe I.
PROCESSAMENTO E APRESENTAÇÃO DO ANTÍGENO
Processamento e apresentação do antígeno são processos que
ocorrem no interior da célula e que resultam na fragmentação de
proteínas (proteólise), associação dos fragmentos com moléculas
do MHC, e expressão das moléculas “peptidio-MHC” na superfície
onde elas poderão ser reconhecidas pelo receptor de célula T na
célula T. Entretanto, a etapa que leva à associação de
fragmentos de proteína com moléculas de MHC diferem no MHC
classe I e classe II. Moléculas de MHC classe I apresentam
produtos de degradação derivados de proteínas intracelulares
(endógenas) no citosol. Moléculas de MHC classe II apresentam
fragmentos derivados de proteínas extracelulares (exógenas) que
estão localizadas em um compartimento intracelular.
Processamento e apresentação do antígeno em células
expressando MHC classe I
Todas as células nucleadas expressam MHC classe I. Como mostrado
na Figura 1, proteínas são fragmentadas no citosol por
proteossomos (um complexo de proteínas com atividade
proteolítica) ou por outras proteases. Os fragmentos são então
transportados através da membrana do retículo endoplasmático por
proteínas de transporte. (As proteínas de transporte e alguns
componentes do proteossomo tem seus genes no complexo MHC). A
síntese e organização das cadeias pesada e beta2
microglobulina ocorre no retículo endoplasmático. No interior do
retículo endoplasmático, a cadeia pesada do MHC classe I, a beta2microglobulina
e o peptídio formam um complexo estável que é transportado à
superfície da célula.
|
|
|
Processamento e apresentação do antígeno em células expressando MHC
classe II
Enquanto
todas as células nucleadas expressam MHC classe I, apenas um limitado
grupo de células expressam MHC classe II, que inclui as células
apresentadoras de antígenos (APC). As principais APCs são macrófagos,
células dendríticas (células de Langerhans), e células B, e a expressão
de moléculas de MHC classe II é tanto constitutiva como induzível,
especialmente pelo interferon-gama no caso dos macrófagos.
Como mostrado na Figura 2, proteínas exógenas incorporadas por
endocitose são fragmentadas por proteases em um endossomo. As cadeias
alfa e beta do MHC classe II, junto com uma cadeia invariante, são
sintetizadas, montadas no retículo endoplasmático e transportadas
através do aparelho de Golgi e trans-Golgi para chegar no endossomo,
onde a cadeia invariante é digerida, e os fragmentos de peptídios da
proteína exógena são capazes de se associar com moléculas de MHC classe
II, que finalmente são transportadas para a superfície da célula.
|
 Figura 2
Figura 2
Via de apresentação de um antígeno exógeno com restrição ao MHC classe
II |
|
|
Outras informações sobre o processamento e apresentação de antígenos
a.
Uma maneira de entender o desenvolvimento de duas vias diferentes é que
cada uma delas finalmente estimula a população de células T que é mais
eficiente na eliminação do antígeno.
Virus
se replicam no interior de células nucleadas no citosol e produzem
antígenos endógenos que podem se associar com MHC classe I. Ao matar
essas células infectadas, células T citolíticas ajudam a controlar a
propagação do virus.
Bacteria reside e se replica principalmente no ambiente extracelular. Ao
ser incorporada e fragmentada no interior de células como antígenos
exógenos que podem se associar com moléculas de MHC classe II, células
auxiliares Th2 podem ser ativadas para ajudar células B a fazerem
anticorpos contra bactéria, o que limita o crescimento desses
organismos.
Algumas bactérias crescem intracelularmente no interior de vesículas de
células como macrófagos. Células T Th1 inflamatórias ajudam a ativar
macrófagos para matar a bactéria intracelular.
b.
Fragmentos de proteínas próprias, assim como de não-próprias se associam
com moléculas de ambas as classes de MHC e são expressas na superfície
da célula.
c. Quais fragmentos se ligam é uma função da natureza química da fenda
para aquela molécula de MHC específica.
|
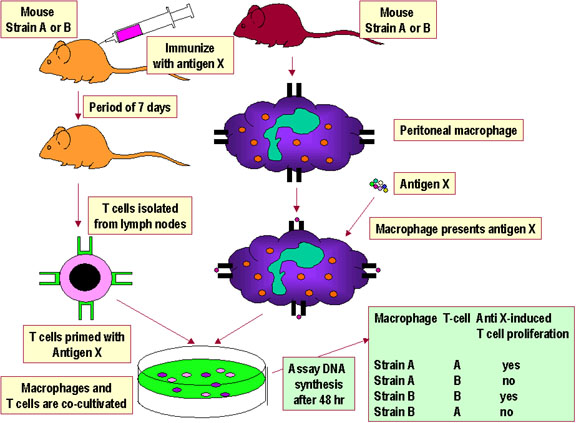
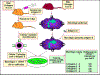 Figura 3
Figura 3
Interações de Restrição do MHC ao próprio de Th/APC.
|
 Figura 4
Figura 4
CTLs virus-específicas de camundongo de uma linhagem A ou linhagem B
lisam apenas células alvo singeneicas infectadas com um virus
específico. As CTLs não lisam células-alvo não infectadas e não são
aloreativas. Análises posteriores mostraram que as CTLs e células alvo
devem ser provenientes de animais que compartilhavam alelos de MHC
classe I para que o alvo apresentasse antígenos virais às CTLs. |
RESTRIÇÃO
DO MHC AO PRÓPRIO
Para que a
célula T reconheça e responda a uma proteína antigênica estranha, ela deve
reconhecer o MHC na célula apresentadora como sendo MHC próprio. Isso é
chamado restrição do MHC ao próprio. Células T auxiliares reconhecem
antígeno no contexto de MHC classe II próprio. Células T citolíticas
reconhecem antígeno no contexto de MHC classe I próprio. O processo pelo
qual células T se tornam restritas ao reconhecimento de moléculas de MHC
próprias ocorre no timo.
Os sistemas experimentais que demonstram a restrição do MHC ao próprio para
interações entre célula APC-T auxiliar e para interações MHC classe I-célula
T citotóxica são mostrados nas Figuras 3 e 4, respectivamente.
|
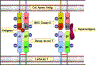
Diferenças entre antígeno e superantígeno. Peptídios antigênicos
são processados no interior da célula e apresentados na superfície da
célula em associação com moléculas do MHC classe II. Eles então
estimulam o receptor de célula T em um linfócito. Superantígenos não são
processados mas eles se ligam à proteína de MHC classe II e à cadeia
beta V do receptor de célula T. Um dado superantígeno ativa uma classe
distinta de células T que expressam uma certa cadeia beta V.
Obs.: No caso da interação MHC II-TCR com um peptídio normalmente
processado, o reconhecimento do peptídio na molécula de MHC requer os
segmentos do TCR V alfa, J alfa, V beta, D beta e J beta. Tal interação
ocorre em baixa frequência. No caso da interação MHC II-TCR com um
superantígeno não processado, apenas uma dada região V beta é
reconhecida. Isso de fato ocorre com uma frequência muito maior. |
CÉLULAS APRESENTADORAS DE
ANTÍGENOS
Os três
tipos principais de células apresentadoras de antígenos são células
dendríticas, macrófagos e células B, embora outras células, que expressem
moléculas de MHC classe II, (e.g., células epiteliais do timo) possam
agir como células apresentadoras de antígenos em alguns casos. Células
dendríticas, que são encontradas na pele e outros tecidos, ingerem antígenos
por pinocitose e transportam antígenos para os linfonodos e baço. Nos
linfonodos e baço elas são encontrados predominantementemente nas áreas de
células T. Células dendríticas são as células apresentadoras de antígenos
mais eficientes e podem apresentar antígenos a células não iniciadas
(virgens). Além disso, elas podem apresentar antígenos internalizados em
associação com moléculas de MHC classe I ou classe II (apresentação
cruzada), embora a via predominante para antígenos internalizados é a via de
classe II. O segundo tipo de célula apresentadora de antígeno é o macrófago.
Essas células ingerem antígenos por fagocitose ou pinocitose. Macrófagos não
são tão eficientes na apresentação de antígenos a células T não iniciadas
mas eles são muito bons na ativação de células T de memória. O terceito tipo
de célula apresentadora de antígeno é a célula B. Essas células se ligam ao
antígeno via sua Ig de superfície e ingere antígenos por pinocitose. Assim
como macrófagos essas célula não são tão eficientes como as células
dendríticas na apresentação de antígeno a células T não iniciadas. Células B
são muito eficientes na apresentação de antígeno a células T de memória,
especialmente quando a concentração de antígeno é baixa devido às Ig de
superfície nas células B se ligarem a antígenos com alta afinidade.
APRESENTAÇÃO DE SUPERANTÍGENOS
Superantígenos são antígenos que ativam células T policlonalmente (ver aula
sobre antígenos) para produzir grandes quantidades de citocinas que podem
ter efeitos patológicos. Esses antígenos devem ser apresentados às células T
em associação com moléculas de MHC classe II mas o antígeno não precisa ser
processado. A figura 5 compara como antígenos convencionais e superantígenos
são apresentados a células T. No caso de um superantígeno a proteína intata
se liga a moléculas do MHC classe II e a uma ou mais regiões Vβ
do TCR. O antígeno não é ligado à fenda de ligação ao peptídio da molécula
de MHC ou à região de ligação ao antígeno do TCR. Assim, qualquer célula T
que usa uma Vβ particular no seu TCR será ativada por um
superantígeno, resultando na ativação de um grande número de células T. Cada
superantígeno se ligará a um conjunto diferentes de regiões Vβ.
|
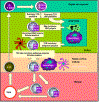
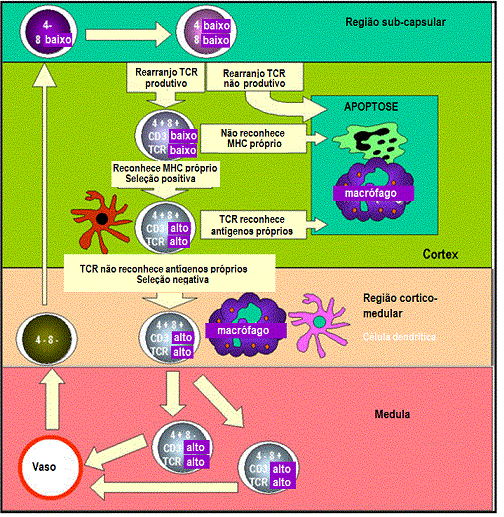
Células pretímicas entram no timo rudimentar e proliferam como grandes
linfoblastos na sua região subcapsular. Os linfoblastos replicam
resultando em um pool de células que se diferenciam. Nesse estágio as
células se tornam CD8 e CD4 positivas mas a expressão é baixa. Genes do
TCR são também rearranjados nessas células e os produtos são também
expressos na superfície da célula em baixos níveis. À medida que as
células maturam, elas movem para o cortex onde aderem a células
epiteliais corticais que são longas e ramificadas, fornecendo uma ampla
superfície para interação com outras células. TCRs nas superfícies dos
timócitos interagem com as moléculas de MHC nas células epiteliais
conduzindo à seleção positiva. As células que não são selecionadas são
sujeitas a apoptose e são fagocitadas pelos macrófagos. À medida que os
timócitos migram avancando em direção ao cortex do timo, a expressão de
CD3, CD4, CD8 e TCR aumenta. TCRs com reatividade ao próprio são
deletados devido ao contato com autoantígenos apresentados pelas células
dendríticas e macrófagos. Isso conduz à seleção negativa. Células que
expressam CD4 ou CD8 aparecem e migram para a periferia
através de vasos especializados na região cortico-medular. |
PAPEL DO TIMO
Tanto
células Th como Tc tem restrição do MHC ao próprio. Além disso, células T
normalmente não reconhecem antígenos próprios. Como são geradas células T
com restrição do MHC ao próprio e por que não são produzidas células T
autorreativas? Rearranjos aleatórios VDJ nas células T poderiam gerar
algumas células T que poderiam reconhecer antígenos próprios. É papel do
timo se certificar de que somente células T que chegam à periferia tenham
restrição do MHC ao próprio e que sejam incapazes de reagir com antígeno
próprio. Células T funcionais na periferia têm que reconhecer antígenos
estranhos associados com MHC próprio, porque células APC ou células alvo
apresentam antígenos estranhos associados com MHC próprio. Entretanto, um
indivíduo não precisa de células T funcionais na periferia que reconheçam
antígenos (próprios ou estranhos) associados com MHC não-próprio. Um
indivíduo especialmente não deseja células T funcionais na periferia que
possam reconhecer antígenos próprios associados com MHC próprio porque eles
poderiam levar a danos em tecidos sadios, normais.
Como resultado de eventos de recombinação aleatória VDJ que ocorrem em
células T imaturas no interior do timo, TCRs de todas as especificidades são
produzidos. Processos no timo determinam quais as especificidades de TCR que
serão mantidas. Há duas etapas sequenciais mostradas na Figura 6. Primeira,
células T com a habilidade de se ligar a moléculas de MHC próprias
expressadas pelas células epitelias corticais do timo são mantidas. Isso é
conhecido como seleção positiva. Aqueles que não se ligam, entram em
apoptose. Assim, células T com a habilidade de se ligar a moléculas de MHC
próprias associadas com moléculas próprias expressadas pelas células
epiteliais do timo, células dendríticas e macrófagos são mortas. Isso é
conhecido como seleção negativa. Aqueles que não se ligam são mantidos. Como
resultado dessas duas etapas, células T tendo um TCR que reconhece MHC
próprio e antígeno estranho sobrevivem. Cada célua T que sobrevive a seleção
positiva e negativa no timo e é liberada na periferia mantém seu receptor de
célula T (TCR) específico.
Enquanto a seleção positiva e negativa está ocorrendo no timo as células T
imaturas estão também expressando antígenos CD4 ou CD8 nas suas superfícies.
Inicialmente a célula pré-T que entra no timo é CD4-CD8-. No timo ela se
torna CD4+CD8+ e à medida que a seleção positiva e negativa se processa a
célula se torna ou uma célula CD4+ ou CD8+. O compromisso de se tornar
células CD4+ ou CD8+ depende de qual seja a classe de molécula de MHC que a
célula encontra. Se uma célula CD4+CD8+ é apresentada com uma molécula de
classe I ela irá regular negativamente CD4 e se tornará uma célula CD8+. Se
a célula é apresentada com uma molécula de MHC de classe II ela irá regular
negativamente CD8 e se tornará uma célula CD4+ (Figura 7).
SELEÇÃO NEGATIVA NA PERIFERIA
A seleção positiva e negativa no timo não é um processo 100% eficiente. Além
disso, nem todos os antígenos próprios são expressados no timo. Assim,
algumas células T autorreativas podem chegar à periferia. Assim, existem
mecanismos adicionais elaborados para eliminar células T autorreativas na
periferia. Esses serão discutidos na aula de tolerância.
Uma vez que células B não têm restrição ao MHC não há necessidade de seleção
positiva de células B. Entretanto, seleção negativa (i.e., eliminação de
clones autorreativos) de células B é necessária. Isso ocorre durante o
desenvolvimento de célula B na medula óssea. Entretanto, seleção negativa de
células B não é crítica como no caso das células T uma vez que, na maioria
das vezes, células B requerem a ajuda de célula T para se tornarem ativadas.
Assim, se uma célula B autorreativa chega à periferia ela não será ativada
devido à falta da ajuda da célula T.
|
| |
 Figura
7 Figura
7
Timócitos precursores CD4- CD8- se tornam células duplo positivas,
expressando baixos níveis de cadeias alfa e beta do receptor de célula T
(TCR). Seleção positiva para a interação com moléculas de MHC-I ou
MHC-II próprias ocorre no epitélio cortical. A maioria das células não
são selecionadas e entram em apoptose. As células que permanecem
inteagem com MHC-I e perdem seus antígenos CD4 ou interagem com MHC-II e
perdem seus antígenos CD8. Células autorreativas são então removidas
como resultado de suas interações com peptídios antigênicos próprios que
são apresentados por células na junção corticomedular com a medula do
timo.
|
| |
|
|
|
 Voltar
à Seção Imunologia do Microbiologia e Imunologia Online
Voltar
à Seção Imunologia do Microbiologia e Imunologia Online
Esta página foi modificada em
Página mantida por
Richard Hunt
Por favor, relate quaisquer problemas para
Richard Hunt
|
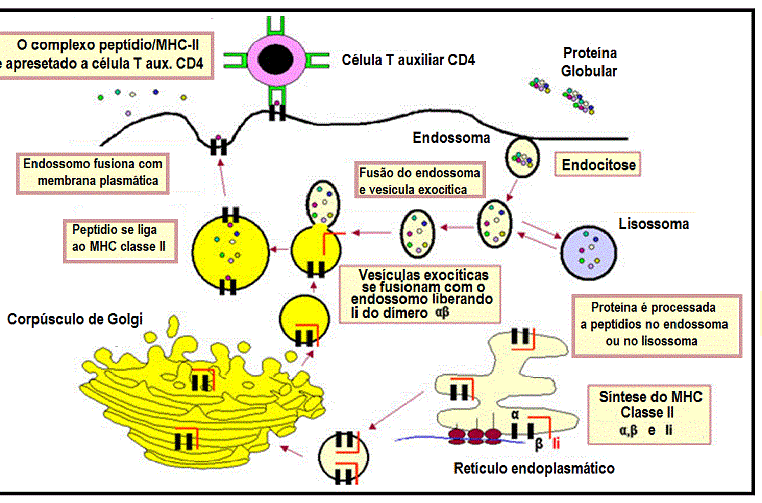 Figura 2
Figura 2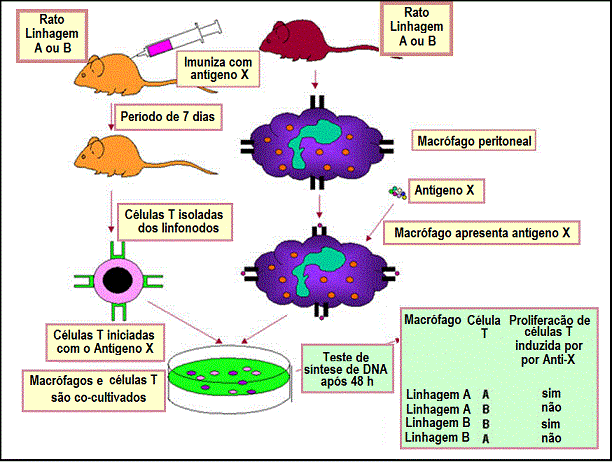 Figura 3
Figura 3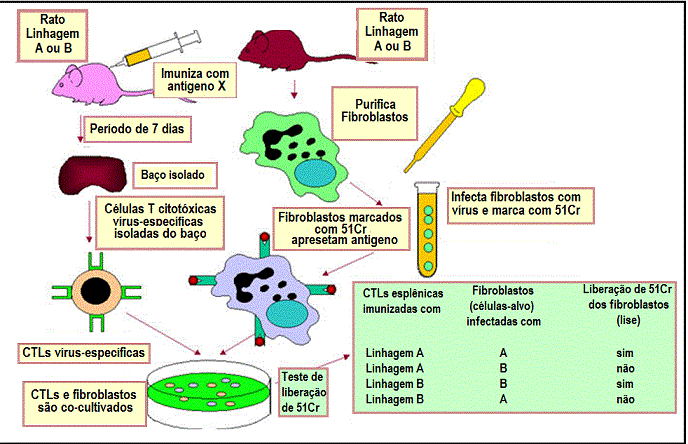 Figura 4
Figura 4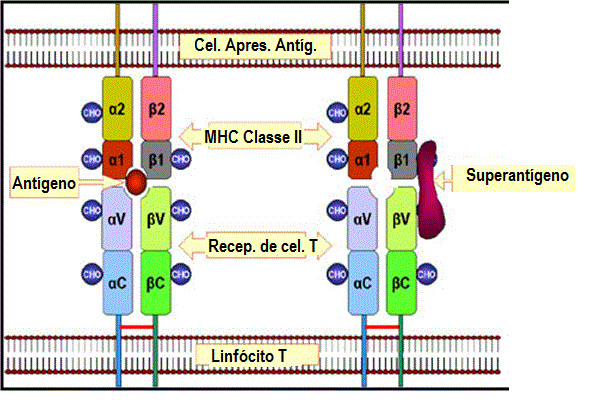
{kind=link}
{kind=link}