|
x |
x |
|
 |
 |
|
INFECTIOUS
DISEASE |
BACTERIOLOGY |
IMMUNOLOGY |
MYCOLOGY |
PARASITOLOGY |
VIROLOGY |
|
VIDEO LECTURE |
IMMUNOLOGIE -
CHAPITRE HUIT
FORMATION DES
ANTICORPS
Gene Mayer,
Ph.D.
Emertius Professor of Pathology, Microbiology and Immunology
University of South Carolina
Denis
Hudrisier, Ph.D.
Centre national de la recherche scientifique (CNRS) · Institute of
Pharmacology and Structural Biology
Université de Toulouse
|
|

 |
|
EN ANGLAIS |
|
EM PORTUGUES - PORTUGUESE |
|
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|
|

 |
|
Logo image
© Jeffrey Nelson, Rush University, Chicago, Illinois and
The MicrobeLibrary |
|
|
|
LECTURES
Male et al. Immunology
7ème édition pp 172-180
|
|
OBJECTIFS DU COURS
Décrire les caractéristiques générales de la réponse immune spécifique
Comparer entre elles les réponses anticorps primaires et secondaires
Décrire les évènements moléculaires mis en jeu lors de commutation de
classe et l’expression membranaire des immunoglobulines
|
CARACTÉRISTIQUES
GÉNÉRALES DE LA RÉPONSE ANTICORPS
Discrimination SOI/NON-SOI
Une des traits caractéristiques de la réponse immunitaire spécifique est
que ce système discrimine normalement les composants du SOI et du NON-SOI
et ne réagit que contre le NON-SOI.
Mémoire
Un second trait de la réponse immune spécifique est qu’elle présente une
mémoire. Le système immunitaire « se souvient » s’il a déjà réagi contre
un antigène auparavant et il réagit alors à une réexposition secondaire
à cet antigène d’une manière différente de celle utilisée lors de la
rencontre initiale dite « primaire ». Généralement, seule une exposition
au même antigène mettra en jeu cette réponse mémoire.
Spécificité
Un troisième caractéristique de la réponse immune spécifique est qu’elle
offre un fort degré de spécificité dans ses réactions. Une réponse
contre un antigène donné sera spécifique à cet antigène ou seulement
étendue à quelques antigènes proches.
N.B. Ces caractéristiques sont propres à toues les
réponses immunes spécifiques.
|
|
MOTS-CLÉS
Phase d’équilibrage
Réponse primaire
Phase de plateau
Commutation de classe
Phase d’induction
Phase de déclin ou de contraction
Phase d’élimination immunitaire
Phase logarithmique
Réponse secondaire ou anamnestique
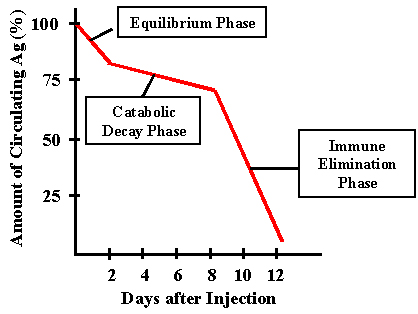
 Figure 1
Figure 1 |
FORMATION DES ANTICORPS
Devenir de l’immunogène
Clairance après une injection primaire
La cinétique de la clairance de l’antigène après une administration
primaire est représentée dans la Figure 1.
Phase d’équilibrage
La première phase est qualifiée d’équilibrage ou encore de phase
d’équilibration. Pendant cette phase, la concentration
antigénique s’équilibre entre les compartiments vasculaires et
extravasculaires par simple diffusion. C’est normalement un
processus rapide. Dans le cas d’antigènes particulaires, comme
ceux-ci ne diffusent pas, cette phase n’existe pas.
Phase de décroissance catabolique
Lors de cette phase, les cellules de l’hôte et les enzymes
métabolisent l’antigène. La majorité de l’antigène est capturé
par les macrophages et les autres cellules phagocytaires. La
durée de cette phase dépend de l’hôte et de l’antigène.
Phase d’élimination immunitaire
Lors de cette phase, les anticorps récemment produits se
combinent à l’antigène pour former des complexes antigène/anticorps
qui sont alors phagocytés et dégradés. Les anticorps
apparaissent dans le sérum seulement à la fin de cette phase
d’élimination immunitaire.
Clairance après une injection secondaire
S’il y a des anticorps circulants dans le sérum, l’injection de
l’antigène pour une seconde fois conduira à une élimination
immunitaire rapide de ce dernier. S’il n’y a pas d’anticorps
circulants, la clairance secondaire de l’antigène passera par les
trois phases indiquées précédemment dans la clairance primaire mais
la phase immunitaire sera opérante plus précocement.
|
 Figure
2
Figure
2 |
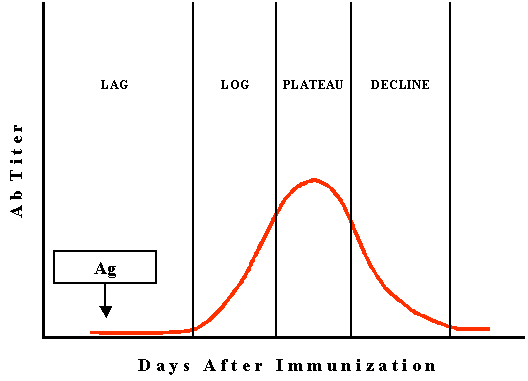
Cinétique de la
réponse anticorps contre un antigène T-dépendant
Réponse anticorps primaire (1o)
La cinétique de la réponse anticorps primaire contre un antigène est
illustrée dans la Figure 2.
Phase d’induction aussi appelée phase de latence
ou retard
Lors de cette phase, l’antigène est reconnu comme étranger et les
cellules B commencent à proliférer et à se différencier en réponse à
la stimulation par l’antigène. La durée de cette phase varie en
fonction de l’antigène mais dure généralement de 5 à 7 jours.
Phase log ou exponentielle
Lors de cette phase, la concentration en anticorps augmente de façon
exponentielle dans la mesure où les cellules B stimulées par
l’antigène se différencient en plasmocytes sécréteurs d’anticorps.
Phase de plateau ou d’équilibre ou stationnaire
Pendant cette phase, il y autant d’anticorps produits que
d’anticorps dégradés de telle sorte que la concentration nette en
anticorps reste constante.
Phase de déclin ou de décroissance (encore
appelée contraction)
Lors de cette phase, le taux de dégradation des anticorps excède
celui de la production de nouveaux anticorps de telle sorte que les
taux globaux d’anticorps diminuent progressivement. A la fin de
cette phase, le taux d’anticorps peut revenir au niveau de base.
|
 Figure
3
Figure
3 |
Réponse
secondaire (2o) encore appelée réponse mémoire ou anamnestique
(Figure 3)
Phase de latence
Lors d’une réponse secondaire, la phase d’induction existe mais est
normalement plus courte que celle observée en réponse primaire.
Phase exponentielle
La phase exponentielle de la réponse secondaire est plus rapide et
conduit à un taux d’anticorps produits plus élevé qu’en phase
primaire.
Phase de plateau
Pendant cette phase, il y autant d’anticorps produits que
d’anticorps dégradés de tel sorte que la concentration nette en
anticorps reste constante.
Phase de contraction
La phase de déclin ou de contraction n’est pas aussi rapide que lors
de la réponse primaire et les anticorps produits peuvent persister
des mois ou des années voire la vie entière.
|
 Figure
4
Figure
4
|
Spécificité des
réponses 1o et 2o
Les anticorps produits en réponse à un antigène sont en
général spécifiques de cet antigène, mais il peut aussi y avoir une
réaction croisée avec d'autres antigènes qui sont similaires sur le plan
structural à l'antigène ayant déclenché la réponse. En général, les
réponses secondaires ne sont provoquées que par le même antigène utilisé
lors de la réponse primaire. Toutefois, dans certains cas, un antigène
apparenté à l’antigène ayant servi à la réponse primaire peut produire
une réponse secondaire, mais c'est une exception rare.
Changements
qualitatifs lors des réponses anticorps 1o et 2o
Changement de classe des
immunoglobulines
Dans la réponse primaire, la classe d'anticorps majoritairement produite
est l’IgM alors que dans la réponse secondaire c’est l’IgG (ou aussi IgA
ou IgE) (Figure 4). Les anticorps qui persistent dans la réponse
secondaire sont les anticorps de type IgG.
|
 Figure 5
Figure 5 |
Affinité
L'affinité des anticorps de type IgG produits augmente
progressivement au cours de la réponse, en particulier après
immunisation avec de faibles doses d'antigène (Figure 5). C’est ce
qu'on appelle la maturation d'affinité. La maturation d'affinité est
plus prononcée lors de la réponse secondaire à l'antigène.
|
 Figure 6
Figure 6 |
Comme illustré dans la Figure 6, la sélection clonale
permet d’expliquer le phénomène de la maturation d'affinité. Une autre
explication à la maturation d'affinité, est que, après qu’un changement
de classe ait eu lieu lors de la réponse immunitaire, des mutations
somatiques ont lieu qui vont modifier les anticorps de telle sorte à ce
qu’ils présentent une plus grande affinité. Il existe des preuves
expérimentales à ce mécanisme, même si on ne connaît pas tous les
détails du mécanisme par lequel la mutation somatique est déclenchée
après l'exposition à l'antigène.
Avidité
En conséquence à l’augmentation de l’affinité au cours de la réponse,
l’avidité des anticorps augmente elle aussi.
Cross-réactivité
L’augmentation progressive de l’affinité au cours de la réponse conduit
également à une augmentation des réactions croisées. Une explication
pour comprendre comment une augmentation d‘affinité conduit à une
augmentation des réactions croisées est fournie dans l’exemple
ci-dessous.
|
| |
|
Affinité de l’anticorps
pour l’antigène (M) |
| |
Précocement |
Tardivement |
| Ag utilisé pour l’immunisation |
10-6
|
10-9
|
|
+ |
++ |
| Ag cross-réactif |
10-3
|
10-6
|
|
- |
+ |
| |
Dans l’exemple montré ici, si une affinité minimale de 10-6 M est
nécessaire pour détecter une réaction, une réaction croisée
d’affinité 10-3 M sera indétectable. Toutefois, si au cours de la
réponse l’affinité augmente d’un facteur 1000, alors la réaction
contre l’antigène ayant servi à l’immunisation mais aussi celle
contre l’antigène cross-réactif seront détectées.
|
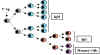 Figure
7
Figure
7 |
Evènements cellulaires au cours des
réponses 1o et 2o contre les antigènes T-dépendants
Réponse primaire (Figure 7)
Phase d’induction ou de latence
Des clones de lymphocytes T et B possédant les récepteurs
antigéniques appropriés se lient à l’antigène, s’activent et
commencent à proliférer. Les clones de lymphocytes B qui se sont
divisés se différencient en plasmocytes qui commencent à secréter
des anticorps.
Phase log ou exponentielle
Au début, le plasmocyte va secréter des IgM car, par défaut, c’est
la région Cμ des gènes de chaînes lourdes qui est associée au exons
VDJ réarrangés, car elle est la plus proche du réarrangement. A la
fin, certaines cellules B feront la commutation de classe d’IgM vers
IgG, IgA ou IgE. Au fur et à mesure que davantage de cellules B
prolifèrent et se différencient en plasmocytes sécréteurs
d’anticorps, la concentration en anticorps augmente de façon
exponentielle.
Phase stationnaire ou de plateau
Au fur et à mesure que l’antigène disparaît, les cellules T et B ne
sont plus activées. De plus, des mécanismes qui régulent
négativement la réponse immunitaire se mettent en place. Finalement,
les plasmocytes commencent à mourir. Quand le taux de production
d’anticorps s’équilibre avec le taux de dégradation des anticorps,
alors la phase stationnaire est atteinte.
Phase de déclin ou de contraction
Quand il n’y a plus d’anticorps produits, l’antigène ayant disparu
et ne pouvant plus activer les cellules T et B, les anticorps
résiduels sont dégradés progressivement conduisant à la phase de
contraction.
|
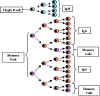 Figure 8
Figure 8
 Figure
9
Figure
9 |
Réponse
secondaire (Figure 8)
Toutes les cellules T et B
stimulées par l'antigène au cours de la réponse primaire ne meurent
pas. Certaines ont une longue durée de vie et constituent ce que
l'on appelle le réservoire de cellules mémoires. A la fois des
cellules T mémoires et des cellules B mémoires sont produites; les
cellules T mémoires survivent plus longtemps que les cellules B
mémoires. Lors d’une nouvelle rencontre avec l’antigène, ce sont
donc non seulement des cellules T et B naïves qui seront activées,
mais aussi les cellules mémoires et, de ce fait, il y a un temps de
latence plus court lors de la réponse secondaire. Comme il y a un
nombre plus important de clone de cellules qui sera stimulé, le taux
de production des anticorps sera également augmenté au cours de la
phase logarithmique de la production d'anticorps et des niveaux plus
élevés de production seront atteints. En outre, étant donné que
beaucoup, si ce n’est la totalité, des cellules B mémoires ont
réalisé une commutation de classe vers IgG (mais aussi IgA ou IgE),
ce sont les IgG qui seront produites précocement lors d’une réponse
secondaire. Finalement, comme il y a aussi davantage de lymphocytes
T mémoires qui peuvent aider les cellules B à faire la commutation
de classe vers IgG (IgA ou IgE), la classe prédominante des
Immunoglobulines produites lors de la réponse secondaire sera l’IgG
(et aussi IgA ou IgE).
Réponse anticorps contre les antigènes T-indépendants
Les réponses immunitaires contre les
antigènes T-indépendants sont caractérisées par la production d'anticorps
IgM et une absence de réponse secondaire. La réponse secondaire contre les
antigènes T-indépendants conduit, en fait, à une nouvelle réponse primaire,
comme illustré dans la Figure 9.
|
 Figure
10
Figure
10 |
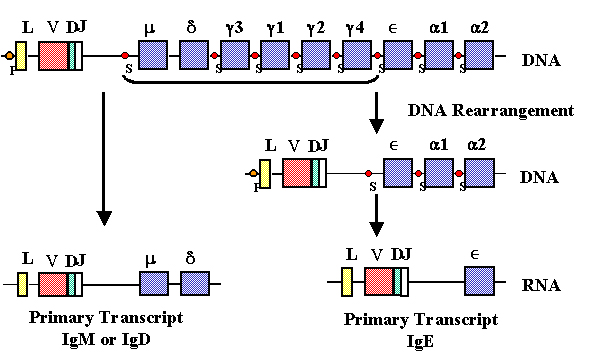
Commutation de classe
Au cours de la réponse immunitaire à un antigène T-dépendant,
une commutation de classe se produit depuis la classe IgM vers une autre
classe (sauf IgD). Notre connaissance de la structure des gènes des
immunoglobulines nous permet de comprendre comment se produit la commutation
de classe (Figure 10).
Au cours de la commutation vers une autre classe d’anticorps,
il se produit un réarrangement d'ADN entre un site de commutation (Su) situé
dans l'intron placé entre les régions VDJ réarrangées et le gène Cμ et un
autre site de commutation situé en amont de l'un des autres gènes de la
région constante de chaîne lourde. Cet événement de recombinaison conduit à
rapprocher la région VDJ de l'un des autres gènes de régions constantes, ce
qui permet l'expression d'une nouvelle classe de chaîne lourde. Comme le
même gène VDJ est amené à proximité d'un gène C différent et que la
spécificité de l'anticorps est déterminée par les régions hypervariables de
la région V, l'anticorps produit après que le commutation ait eu lieu aura
la même spécificité que précédemment.
Ce sont les cytokines sécrétées par les cellules T
auxiliaires qui vont provoquer la commutation vers certains isotypes
d’anticorps.
|
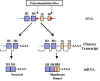 Figure 11
Figure 11 |
Immunoglobuline
membranaire et sécrétée
La spécificité des immunoglobulines de membrane exprimées à
la surface d’une cellule B et de celle de l'Ig secrétée par la forme
plasmocytaire de cette cellule B est la même. Les raisons expliquant
comment, à partir d'une cellule B donnée, la spécificité de
l’immunoglobuline présente à la membrane sera identique à celle sécrétée
peuvent être déduites de l’organisation des gènes d'immunoglobulines (Figure
11).
Il y a deux sites potentiels de polyadénylation du gène de
l'immunoglobuline. L'un est situé après l'exon codant pour le dernier
domaine de la chaîne lourde, et l'autre, après les exons qui codent pour les
domaines trans-membranaires. Si le premier site polyA est utilisé, le
pré-ARNm sera épissé pour produire une protéine sécrétée. Si le second site
polyA est utilisé, le pré-ARNm se épissé pour produire une forme membranaire
de l'immunoglobuline. Cependant, dans tous les cas, la région VDJ utilisée
est la même et, donc, la spécificité de l'anticorps restera la même. Tous
les gènes de régions C ont ces portions supplémentaires codant pour des
parties ancrées dans la membrane, et donc, après la commutation vers
d'autres classes d'immunoglobulines, celles-ci peuvent être soit sécrétées
soit exprimées à la surface des cellules B.
|
|
|
 Retourner à la section d'immunologie de Microbiologie et Immunologie On-line
Retourner à la section d'immunologie de Microbiologie et Immunologie On-line
This page last changed on
Friday, December 07, 2012
Page maintained by
Richard Hunt
Please report any problems to
richard.hunt@uscmed.sc.edu
|
 Figure
2
Figure
2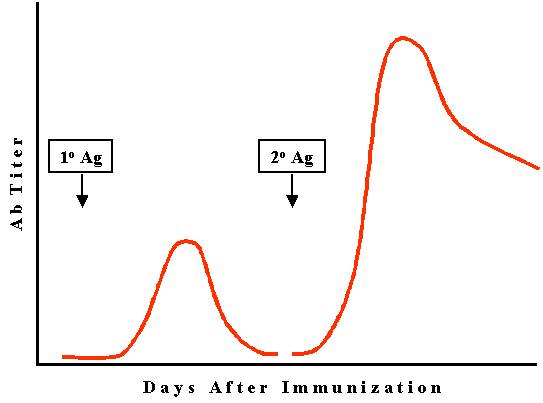 Figure
3
Figure
3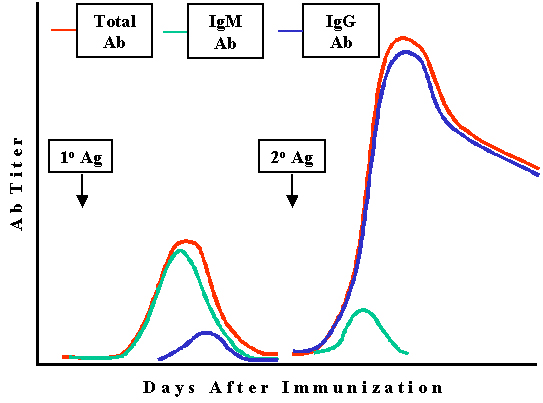 Figure
4
Figure
4
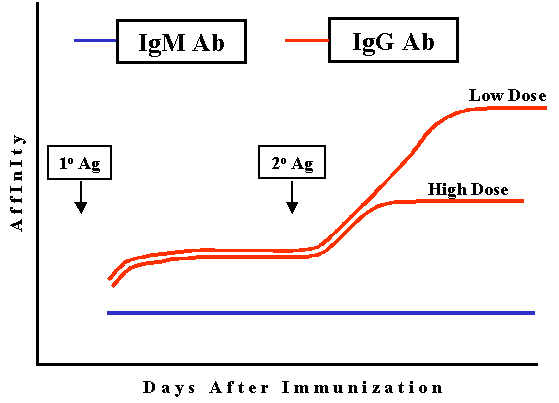 Figure 5
Figure 5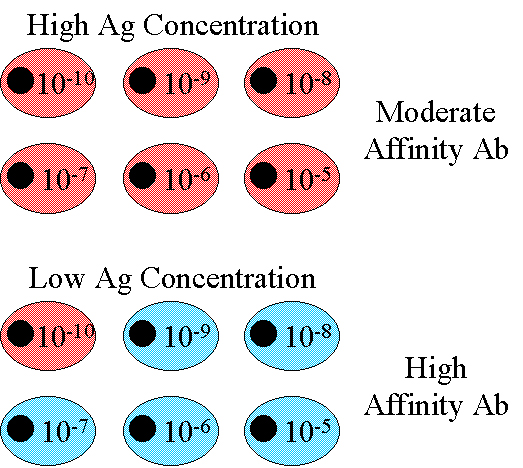 Figure 6
Figure 6 Figure
7
Figure
7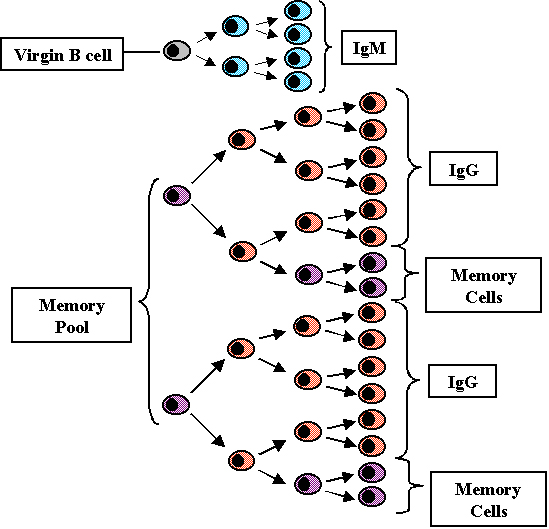 Figure 8
Figure 8
 Figure
10
Figure
10 Figure 11
Figure 11