| x | x | |||||||||||||||||||||||||||||||
|
|
 |
|||||||||||||||||||||||||||||||
| BAKTERIYOLOJİ | IMMUNOLOGY | MYCOLOGY | PARASITOLOGY | VIROLOGY | ||||||||||||||||||||||||||||
|
|
|
|||||||||||||||||||||||||||||||
| ENGLISH | ||||||||||||||||||||||||||||||||
| SPANISH | ||||||||||||||||||||||||||||||||
| PORTUGUESE | ||||||||||||||||||||||||||||||||
| SHQIP - ALBANIAN | ||||||||||||||||||||||||||||||||
| FARSI | ||||||||||||||||||||||||||||||||
|
Let us know what you think |
||||||||||||||||||||||||||||||||
| SEARCH | ||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
| Logo image © Jeffrey Nelson, Rush University, Chicago, Illinois and The MicrobeLibrary | ||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
|
ANAHTAR KELİMELER |
Bakteriler, büyüme gereksinimleri için enerji kaynağı olarak, "organik" karbon (örneğin, şeker ve yağ asitleri) ve metal iyonları (örneğin, demir) kullanmaktadır. En uygun sıcaklık, pH ve oksijen gereksinimi (veya oksijen ihtiyacının olmaması) önemlidir. Oksijen Gereksinimleri Zorunlu aerobik bakterilerin büyümesi oksijen varlığında olur; onlar fermantasyon yapamazlar. Zorunlu anaeroblar oksidatif fosforilasyon yapamazlar. Ayrıca bunlar oksijen tarafından öldürülür; bunlar örneğin, katalaz [hidrojen peroksiti, H2O2, su ve oksijene dönüştürür]. peroksidaz [enzim tarafından NADH + H2O2, NAD ve O2'ye dönüştürülür] ve süperoksit dismutaz [enzim tarafından süperoksit O2, H2O2 dönüştürülür] gibi bazı enzimlerden yoksundur. Bu enzimler, oksijen varlığında metabolizma süresince üretilen peroksit ve oksijen gibi serbest radikalleri zararsız hale getirirler. Aerotoleran anaeroblar anaerobik teneffüs eden bakterilerdir, ancak oksijen varlığında yaşayamazlar. Fakültatif anaeroblar hem fermantasyon hem de aerobik solunum gerçekleştirebilirler. Bu bakterilerde oksijen varlığında anaerobik solunum genellikle kapatılır ve bu organizmalar aerobik teneffüsle devam ederler. Mikroaerofilik bakteriler düşük oksijen konsantrasyonlarında iyi büyürler fakat bu bakteriler için yüksek oksijen konsantrasyonları öldürücü olabilir. Besin Gereksinimleri Besin kaynakları organik karbon, azot, fosfor, kükürt ve demir gibi metal iyonlarını içerir. Bakteriler demir bağlayan küçük moleküller salgılarlar (sideroforlar, örneğin; enterobaktin, mikobaktin). Siderofor (Bağlı demir) bakteri hücresi tarafından reseptörleri aracılığıyla tekrar geri alınır. İnsanda demir taşıma proteinleri (örneğin, transferin) vardır. Bundan dolayı bakteriler demir için ev sahibi ile rekabet açısından kötü patojenlerdir. Sıcaklık Bakteriler suyun donma ısısına yakın derecelerden, kaynama noktasına yakın derecelere kadar çeşitli derecelerde gelişebilir. Bu aralığın ortasında iyi büyüyenler mezofilik olarak adlandırılırlar ve bu türler tüm insan patojenleri ve oportünistleri içerirler. (Sırasıyla, optimum düşük ısıda ve yüksek sıcaklıkta üreyen bakteriler psikrofiller ve termofiller olarak da bilinir). pH Birçok bakteri nötr pH'da en iyi büyürler. Ancak bazı bakteriler tamamen asit
veya alkali koşullar altında hayatta kalabilirler ve hatta büyüyebilirler. |
|||||||||||||||||||||||||||||||
|
FİLM (movie) |
||||||||||||||||||||||||||||||||
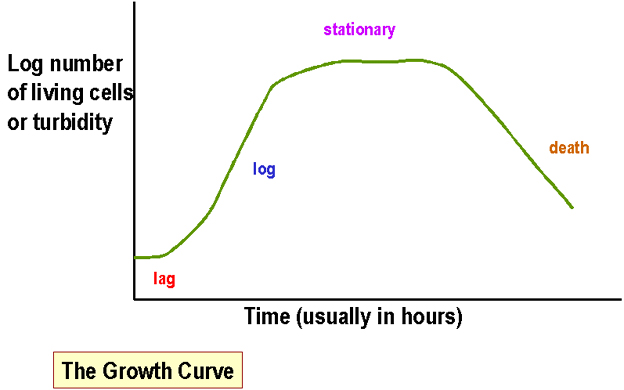 Şekil
1. Büyüme Eğrisi Şekil
1. Büyüme Eğrisi |
Bakterilerin sıvı kültürlerde kütle ölçümü
|
|||||||||||||||||||||||||||||||
|
|
ŞEKERİN METABOLİZMASI (metabolik yolların
bir örneği)
* Moleküldeki toplam karbon sayısı anlamına gelir
Şekerin katabolize edilmesinde, ATP içinde depolanmış enerji üretmek için, bakterilerde bu yola alternatif yollar vardır. Birisi hayvan ve bitki hücrelerinde bulunan pentoz fosfat yolu (heksoz monofosfat şantı) dır. NADPH bu yol kullanılarak oluşturulur. Diğer bir katabolizma yolu, Entner Doudoroff yolu, genel olarak sadece belirli bakteri hücrelerinde bulunur. Anaerobik Solunum Anaerobik solunum glikoliz ve fermentasyonu içerir. Bu işlemin sonraki
safhalarında, (glikoliz sırasında üretilen) NADH bir hidrojen kaybederek
NAD'e geri dönüştürülür. Hidrojen piruvata aktarılır ve bakteri türüne bağlı
olarak çeşitli metabolik son ürünler üretilir.
Aerobik solunum Aerobik solunum glikolizi ve trikarboksilik asit döngüsünü (Krebs döngüsü) içerir. Piruvat tamamen yıkılarak karbon dioksite (C1) dönüştürülür ve bu süreçte NAD NADH'a dönüştürülür. Bu nedenle, aerobik fermantasyonda, NADH iki kaynaktan (glikoliz ve Krebs döngüsü) oluşturulur. Oksidatif fosforilasyonda NADH'ın NAD'a geri dönüşümü aşırıdır ve süreçte daha fazla ATP (depolanan enerji) üretilir. Ubikinonlar ve sitokromlar bu son süreçte yer alan elektron taşıma zincirinin bileşenleridir. Oksijenin suya dönüşümü sürecin son adımıdır.
|
|||||||||||||||||||||||||||||||
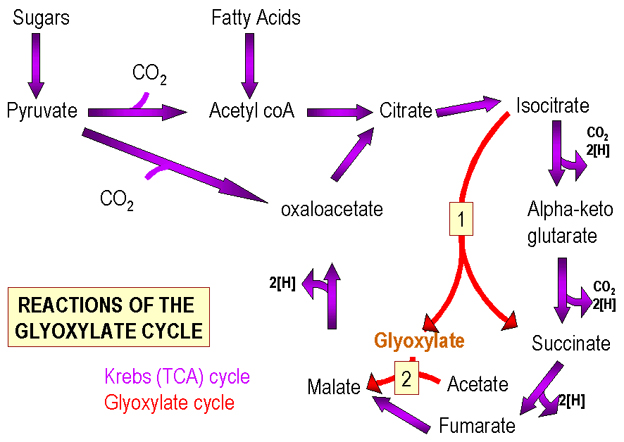 Şekil 2. Krebs ve Glioksilat Döngüleri
Şekil 2. Krebs ve Glioksilat Döngüleri |
Oksidatif fosforilasyon
a) Piruvattan asetil CoA'ya CO2 kaybını (C1) döngü bileşenine bir C4 ilavesi
izler, (oksaloasetat) bir C6 bileşen (sitrik asit) üretir. Bu şekilde üretilen
C6 molekül sayısı başlangıçta mevcut C4 moleküllerinin sayısına eşittir.
b) Piruvata CO2 ilave edilmesi ile bir C4-bileşiğini üretilir. Bu durumda, C4'ün ek molekülleri (bir döngü bileşeni) oluşturulur.
Dolayısıyla eğer döngü bileşenlerin bazıları diğer biyosentetik yollarda kullanılmak üzere taşınırsa, bu reaksiyon ile yeri yeniden doldurulabilir.
|
|||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
|
|
Yağ asitlerinin metabolizması
Yağ asitleri asetil gruplara (C2) yıkılır ki, bu gruplar Krebs döngüsü
besleyerek bir C4' ara bileşen ilavesiyle bir C6 üretimini sağlar. Döngü
sırasında, eklene C2 CO2 olarak kaybolur ve C4 yeniden oluşturulur. Genel olarak,
döngünün ara moleküllerin sayısındaki bir artış meydana gelmez. Bu yüzden, yağ
asitler, tek karbon kaynağı ise hiçbir Krebs döngüsü ara ürün olarak kapatmadan
çıkarılabilir:
Krebs döngüsü yerine, bakteri glioksilat çevrimi kullanabilir, (Şekil 2) (modifiye bir Krebs Döngüsü) hangi enzimatik adımlar içinde hangi iki CO2 molekülü C6 ara ürününden by-pass edilerek çıkarılır. C6-ara ürünü iki C4 bileşiğine dönüştürülür (döngünün her iki bileşeni). Bu durumda, (yağ asitlerinden) her asetil grubu için, bir ek döngü ara ürünü üretilebilir. Glioksalat geçiş yolu genellikle hayvan hücrelerinde bulunmadığı için gıda da hazır bulunan, önceden oluşturulmuş yağ asitleri kullanılır.
Özetle, Krebs döngüsü bir biyosentez ve enerji üretim görevi yapar. Bununla birlikte, ara ürünler başka metabolik yollarda kullanılmak için çıkarılması durumunda, bunların yeri tekrar doldurulmalıdır. Yenileme işlemi şeker ve yağ asitlerinin kullanımında farklıdır. |
|||||||||||||||||||||||||||||||
|
|
|
|||||||||||||||||||||||||||||||