|
x |
x |
|
 |
 |
|
INFECTIOUS
DISEASE |
BAKTERIYOLOJİ |
IMMUNOLOGY |
MYCOLOGY |
PARASITOLOGY |
VIROLOGY |
|
|
BAKTERİYOLOJİ – BÖLÜM
DOKUZ
GENETİK DÜZENLEME MEKANİZMALARI
Gene Mayer, PhD
Emeritus Professor
University of South Carolina School of Medicine
Çeviren.
Prof. Dr. Mustafa Demirci
İzmir Katip Çelebi Üniversitesi,
Tıp Fakültesi, Tıbbi Mikrobiyoloji A.D. İzmir, Türkiye
|
|
TURKISH |
|
SPANISH |
|
ALBANIAN |
|
|
Let us know what you think
FEEDBACK |
|
SEARCH |

 |
|
Logo image © Jeffrey
Nelson, Rush University, Chicago, Illinois and
The MicrobeLibrary |
|
ÖĞRENİM HEDEFLERİ
Bakteri genlerinin yapısı ve transkripsiyonu tartışmak
Bakterilerde moleküler mekanizmaları tanımlamak için gen aktivitesininin
düzenlenmesini kullanmak
Uyarılabilir ve bastırılabilir operonlan karşılaştırmak ve zıtlıklarını
tanımlamak
Katabolik baskılara ve zayıflamada rol oynayan moleküler mekanizmaları
tanımlamak Bakterilerde enzim aktivitesini düzenleyen yolları tartışmak
|
GEN EKSPRESYONUNUN DÜZENLENMESİ
Bakteriler, her zaman bütün yapabildikleri proteinleri yapmazlar. Aksine,
onlar çevreye uyum sağlayabilirler ve sadece belirli bir ortamda hayatta kalmak
için gerekli olan gen ürünlerini yapabilirler. Örneğin, bakteriler ortamda bol
triptofan kaynağı olduğunda triptofan yapmak için gerekli enzimleri sentez
etmezler. Triptofan ortamda bulunmadığı zaman bu enzimler yapılır. Benzer
şekilde, bir bakteride bir antibiyotiğe direnç kazandıran bir gen bulunduğu
zaman bu genin eksprese edileceği anlamına gelmez. Direnç geni, sadece
antibiyotik ortamda mevcut olduğunda ifade edilebilir.
Bakteriler genellikle, gen ekspresyonunu mRNA transkripsiyonu seviyesinde
düzenleyerek kontrol etmektedir. Bakterilerde bu fonksiyonu gören genler
genellikle birbirlerine bitişik yer alır ve koordineli düzenlenir (örn; biri
eksprese edildiğinde, hepsi eksprese edilir). Kümelenmiş genlerin koordineli
düzenlenmesi bir polisistronikmRNAüretimininin düzenlenmesi ile gerçekleştirilir
(örn; birkaç genin bilgilerini içeren geniş bir mRNA). Böylece, bakteriler
çevrelerine "duyarlı" dırlar ve ortam için gerekli olan genlerin
transkripsiyonunu düzenleyerek uygun bir dizi eksprese ederler.
|
|
ANAHTAR KELİMELER
Gen ekspresyonunu koordine etmek, Polisistronik m-RNA, Promoter (başlatıcı),
Operon, İndüklenebilir operon, İndükleyici, Yapısal gen, Düzenleyici
gen, Represör, Operator, Negatif kontrol, Katabolit baskısı, CAP
proteini, Pozitif kontrol, Bastırılabilir operon, Co-represör, Apo-represör,
Azaltma-zayıflama, Lider bölgesi, Geri Bildirim inhibisyonu, Epigenetik
değişiklik.
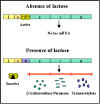
 Şekil 1; Laktoz operon
Şekil 1; Laktoz operon
 Şekil 2; Glukozun varlığında veya yokluğunda lak genlerinin
transkripsiyonu.
Şekil 2; Glukozun varlığında veya yokluğunda lak genlerinin
transkripsiyonu. |
Indüklenebilir genler -
operon modeli
Tanım
Bir uyarılabilir gen ortamında bir madde (bir
indükleyici) varlığında eksprese edilen bir gendir. Bu madde, bu
maddenin metabolizmasında rol oynayan bir ya da daha fazla genin (yapısal
genler) ekspresyonunu kontrol edebilirler. Örneğin, laktoz
metabolizmasından sorumlu olan, lak genlerin ekspresyonuna laktoz neden
olur. Bir antibiyotik profilaksi bu antibiyotiğe karşı dirence yol açan
bir genin ekspesyonuna neden olabilir.
ndüksiyon, bir maddenin normal yolda indükleyici olduğu
ve bu maddenin katabolizmasıyla sonuçlanan yaygın metabolik yoldur.
Laktoz Operon
Yapısal genler
Laktoz operonu (Şekil 1), laktoz metabolizmasında rol oynayan
enzimleri kodlayan üç yapısal geni içerir.
-
Lac z geninin kodladığı Β-galaktosidaz, laktozu
glukoz ve galaktoza parçalayan bir enzimdir.
-
Lac y geninin kodladığı permeaz, laktoz alımında
görev alan alır.
-
Lak a geni galaktoztransacetylase'ı kodlar.
Bu genler, üç enzim vermek üzere kopyalanan bir
polisistronikmRNA'ya bir ortak başlatıcıdan transkripte edilir.
Düzenleyici gen
Yapısal genlerin sentezlenmesi, sadece uyarıcının varlığında veya
yokluğunda etkilenmez, aynı zamanda belirli bir düzenleyici gen
tarafından kontrol edilir. Düzenleyici gen etkilediği genin
bitişiğinde veya uzağında olabilir. Düzenleyici gen, represör olarak
adlandırılan belirli bir protein ürünü kodlar.
Operatör
Represör, operatör olarak adlandırılan ve regüle edilen yapısal
genlere bitişik olan DNA'nın belirli bir bölgesine bağlanarak etki
eder. Öperatör bölgesi ve promotör ile birlikte yapısal genler bir
operon olarak adlandırılır. Bununla birlikte, represörün operatöre
bağlanması indükleyici ile önlenir ve indükleyici de zaten operatöre
bağlı olan represörü uzaklaştırır. Böylece, indükleyicinin
varlığında represörinaktifdir, operatöre bağlanmaz ve süreç yapısal
genlerin transkripsiyonu ile sonuçlanır. Bunun aksine, uyarıcının
yokluğunda represöraktifdir, operatöre bağlanır ve yapısal genlerin
transkripsiyonun inhibisyonu ile sonuçlanır. Yapısal genlerinin
transkripsiyonu kapatılması düzenleyici gen ürünlerinin (represör)
fonksiyonu olduğu için bu tür kontrol, NEGATİF KONTROL olarak
adlandırılır.
İndükleyici
Lak genlerinin transkripsiyonu bir indükleyici (laktoz veya diğer β-galaktozitler)
varlığında veya yokluğunda etkilenir (Şekil 2).
|
Örn, |
+ indükleyici |
Ekspresyon |
|
- indükleyici |
Ekspresyon
yok |
|
|
|
 Şekil 3;Katabolit baskılama
Şekil 3;Katabolit baskılama
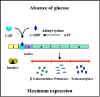 Şekil 4; Lacoperon tarafından kodlanan
proteinlerin ekspresyonu üzerindeki glukozun etkisi
Şekil 4; Lacoperon tarafından kodlanan
proteinlerin ekspresyonu üzerindeki glukozun etkisi
 Şekil 5; Lacoperon tarafından kodlanan proteinlerin ekspresyonu
üzerindeki glukozun etkisi
Şekil 5; Lacoperon tarafından kodlanan proteinlerin ekspresyonu
üzerindeki glukozun etkisi |
Katabolit baskılama (represyon) (Glikoz
Etkisi) Birçok indüklenebilir operonlar sadece ilgili
indükleyicileri ve düzenleyici genler tarafından kontrol edilmez,
aynı zamanda ortamdaki glikoz seviyesiyle de kontrol
edilmektedir.Glukozun, bir dizi farklı indüklenebilir operonların
ekspresyonunu kontrol etme, yeteneği katabolit baskılama olarak
adlandırılır. Bu durum Şekil 3'te gösterilmiştir.
Katabolit baskılama genellikle, bir enerji kaynağı olarak
kullanılan bileşiklerin parçalanmasında görevli olan operonlarda
görülür. Glukoz bakteriler tarafından tercih edilen enerji kaynağı
olduğu için, diğer operonların ekspresyonunun regüle edilmesinde
glukozun yeteneği, enerji kaynağı olarak herhangi bir karbon
kaynağından önce bakterilerin glikozdan yararlanmasını garanti eder.
Mekanizma
Bakterilerdeki glukoz ve siklik AMP (cAMP) seviyeleri arasında
ters bir ilişki bulunmaktadır. Glukoz düzeyleri yüksek olduğunda
cAMP düzeyleri düşüktür ve glikoz düzeyleri düşük olduğunda cAMP
düzeyleri yüksektir. Bu ilişki, glukozu hücre içerisine
taşınması durumunda cAMP üreten enzim olan adenilsiklazıinhibe
etmesi temelindedir. Bakteri hücresinde cAMP, CAP veya CRP
olarak adlandırılan bir cAMP bağlayıcı proteine bağlanır. cAMP-CAP
kompleksi, serbest CAP proteini değil, katabolit baskılamaya
duyarlı operonların başlatıcı bölgesinde bir yere bağlanır.
Şekil 4 ve 5'de gösterildiği gibi, kopleksin bağlanması daha
etkin bir promotör ve böylece promotorden (başlatıcı) daha etkin
transkripsiyonların başlaması ile sonuçlanır. CAP-cAMPkompleksinin
rolü transkripsiyonu açmak için olduğundan bu tip kontrol
mekanizması POZİTİF KONTROL olarak adlandırılır. Bu tür
kontrolün sonuçları olarak, bir katabolit baskılamaya duyarlı
operonun maksimum eksresyonunun olması için, glukoz ortamda
mevcut olmalı ve operonun indükleyicisinin de olması gerekir.
Her ikisi de olduğu takdirde, glikoz metabolize olana kadar,
operon maksimum eksprese edilmez. Açıkçası,indükleyici mevcut
olmadıkça operonun hiçbir ekspresyonu olmayacaktır.
|
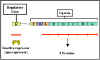 Şekil 6; Triptofan operon
Şekil 6; Triptofan operon
 Şekil 7; Tryp operondan ekspresyonu üzerine triptofan etkisi
Şekil 7; Tryp operondan ekspresyonu üzerine triptofan etkisi |
Bastırılabilir genler - operon modeli
Tanım
Baskılanabilir genler, ortamında bir madde (bir yardımcı-represör)
varlığında, bu maddenin metabolizmasında rol alan genlerin (yapısal
genler) ekspresyonunu kapatan genlerdir.
örneğin, TriptofanTrp genlerin ekspresyonunu bastırmıştır.
Baskılama metabolik yollar içinde; bir maddenin biyosentezinde ve
genelde yolun son ürünü olarak regüle edilen co reseptör olarak,
yaygındır.
Triptofanoperon
Yapısal genler
Triptofanoperon (Şekil 6), triptofan sentezinde yer alan enzimler
için, beş yapısal genleri içerir. Bu genler, beş enzimi sentezletmek
üzere, bir polisistronikmRNA'ya bir ortak promotörden transkripte
edilir.
Düzenleyici gen
Yapısal genlerin ekspresyonu, sadece ko-represör varlığında veya
yokluğunda etkilenmez, aynı zamanda belirli bir düzenleyici gen
tarafından da kontrol edilir. Düzenleyici gen etkilediği genin
bitişiğinde ve uzağında olabilir. Düzenleyici gen, REPRESÖR olarak
adlandırılan, spesifik bir proteini kodlar (bazen apo-represör
olarak adlandırılır). Represör sentezlendiği zaman aktif değildir.
Bununla birlikte, (örneğin, triptofan) ko-represör ile birleşerek
aktif hale getirilebilir.
Operator
Aktif represör / ko-represör kompleks, operatör olarak adlandırılan
ve düzenlenen yapısal gene bitişik olan DNA'nın spesifik bir
bölgesine bağlanarak etki eder. Operatör bölgesi ve promotörü ile
birlikte yapısal genler bir operon olarak adlandırılır. Bu nedenle,
ko-represör varlığında represör aktiftir ve operatöre bağlanır ki bu
durum yapısal genin transkripsiyonunun baskılanmasıyla sonuçlanır.
Bunun aksine, ko-represör yokluğunda represör inaktiftir ve
operatöre bağlanmaz ki bu durum yapısal genlerin transkripsiyonu ile
sonuçlanmaz. Düzenleyici gen ürününün (represör) fonksiyonu, yapısal
genin transkripsiyonunu kapatmak olduğu için, bu tür kontrol bir
NEGATİF KONTROL olarak adlandırılır.
Ko-represör
Triptofan gen transkripsiyonu, bir ko-represör (triptofan) (Şekil 7)
varlığında veya yokluğunda etkilenir.
| e.g.
|
+ ko-represör |
ekspresyon yok |
|
- ko-represör |
ekspresyon |
|
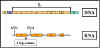 Şekil 8; Atenüasyonun mekanizması
Şekil 8; Atenüasyonun mekanizması
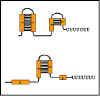 Figure 9 Formation of stem-loops
Figure 9 Formation of stem-loops |
Atenuasyon (Sönme)
Birçok bastırılabilir operonlarda, promotör de başlatılan transkripsiyon
ilk yapısal gen öncesinde bulunan bir lider bölgede prematür olarak
kesilebilir. (örn.; polimeraz transkripsiyonu sona erdirmeden önce ilk geni
operona alır). Transkripsiyonun erken sonlandırıldığı bu durum; ATENUASYON (SÖNME)
olarak adlandırılır. Her ne kadar sönme belirli sayıda operonda görülse de,
mekanizması en iyi amino asit biyosentezine katılan baskılanabilir
operonlarda anlaşılmıştır. Bu durumlarda sönme akraba aminoaçil t-RNA
varlığı ile düzenlenir.
Mekanizma (Bakınız; Şekil 8)
Transkripsiyon promoterde başlatıldığı zaman, aslında önce birinci
yapısal genin transkripsiyonu başlar ve lider transkript yapılır. Bu
lider bölgesi, protein sentezi için, başlangıç ve durdurma sinyali
içerir. Bakteriler bir nükleer membrana sahip olmadıkları için,
transkripsiyonu ve translasyon aynı anda oluşabilir. RNA polimerazın
lider bölge transkripsiyonu sırasında kısa bir peptid yapılır. Bu test
peptit molekülünün ortasında birkaç triptofan kalıntısı bulunur. Bu
nedenle, eğer bir test peptidi translasyonu için triptofanil-t-RNA
yeterli bir miktarda mevcutsa tüm peptid yapılacak ve sonra ribozoma
durdurma sinyali ulaşacaktır. Öte yandan, bu peptid translasyonu için
yeterli triptofanil-t-RNA yok ise ribozom durdurma sinyalini almadan
önce iki triptofan kodonunda durdurulacaktır.
Lider m-RNA dizisi tamamlayıcı dizileri olan (Şekil 9) dört bölge
içerir. Bu nedenle, çok sayıda farklı ikinci kök ve halka yapıları
oluşturulabilmektedir. Bölge 1 yalnızca bölge 2 ile baz çiftleri
oluşturabilir; Bölge 2, bölge 1 veya 3 ile baz çiftleri oluşturabilir;
bölge 3, bölge 2 veya 4 ile baz çiftleri oluşturabilir; ve 4 bölgesi,
sadece bölge 3 ile baz çifti oluşturabilir. Böylece RNA'da üç olası kök
/ halka yapısı oluşturulabilir.
bölge 1 : bölge 2
bölge 2 : bölge 3
bölge 3 : bölge 4
Olası yapılardan biri, (bölge 3 ile bölge 4'ün baz çifti yapması) RNA
polimerazın transkripsiyonu sona erdirmesi için, bir sinyal üretir (örn;
transkripsiyonun atenüasyonu). Bununla birlikte, tek bir gövde ve halka
yapısının (baz çifti) oluşumu diğer oluşumları engeller. Eğer bölge 1
ile bölge 2 baz çifti oluşturur ise bölge 2'nin bölge 3 ile baz çifti
oluşturması mümkün değildir. Benzer şekilde eğer bölge 3 bölge 2 ile baz
çifti oluşturursa bölge 4 ile bölge 3'ün baz çifti yapması imkansız olur.
Çeşitli kök ve halka yapılarının oluşumunu etkileyecek test peptidi
translasyonu için ribozomların yeteneği Şekil 10. Ribozoma translasyon
için durdurma sinyali ulaşırsa, ribozom bölge 2 kapsayacak ve böylece
bölge 2'nin diğer bölgeler ile baz çifti oluşturması mümkün olmayacaktır.
Bu durum transkripsiyonun sonlandırılması sinyali üretimine yol açacak
olan, bölge 3 ile bölge 4'ün eşleşmesini mümkün hale getirecektir. Bu
nedenle, test peptidi translasyonu için yeterli triptofanil-t-RNA
olduğunda, atenüasyon ortaya çıkar ve yapısal genlerin transkripsiyonu
olmaz. Bunun aksine, test peptidi translasyonu için triptofanil-t-RNA
yetersiz olduğu zaman herhangi bir atenüasyon olmayacaktır. Ribozom
bölge 1'de iki triptofan kodonunda durması nedeniyle bölge2'nin bölge 3
ile baz çifti oluşturmasına imkan olur ve atenüasyon sinyalinin oluşması
önlenir (örn.; bölge 3 bölge 4 ile baz çifti yapmıştır). Bu nedenle,
yapısal gen transkribe edilir.
|
 Şekil 10; Atenüasyon mekanizması
Şekil 10; Atenüasyon mekanizması |
| |
ENZİM AKTİVİTESİNİN REGÜLASYONU
Bakteriler de kendi enzimlerin aktivitelerini düzenleyen yollara sahiptirler.
Geribildirim inhibisyonu
Bakteriyel enzim aktivitesi, genellikle geribildirim inhibisyonuna
tabidir. Genellikle, bir yolun son ürünü inhibitördür ve yoldaki ilk enzimde
düzenleme basamağıdır.
Epigenetik modifikasyon
Bakteriyel enzimlerin aktivitesi bu enzimlerin kovalent modifikasyonları
ile kontrol edilebilir. Bu tür modifikasyon EPİGENETİK MODİFİKSYON olarak
adlandırılır.
örn.;Glutamin sentetazın adenilasyonu
Glikojen sentetazın fosforilasyonu
Genellikle bu değişiklikler, enzimlerin aktivitelerinin açılıp ve kapana
bilmesi için geri dönüşümlüdür.
|
|
|
 Mikrobiyoloji ve İmmünoloji On-line Kitabı Bakteriyoloji Bölümüne dön
Mikrobiyoloji ve İmmünoloji On-line Kitabı Bakteriyoloji Bölümüne dön
This page last changed on
Wednesday, March 02, 2016
Page maintained by
Richard Hunt
|
 Şekil 3;Katabolit baskılama
Şekil 3;Katabolit baskılama Şekil 6; Triptofan operon
Şekil 6; Triptofan operon Şekil 10; Atenüasyon mekanizması
Şekil 10; Atenüasyon mekanizması




