|
xx |
x |
|
 |
 |
|
INFECTIOUS
DISEASE |
BAKTERIYOLOJİ |
IMMUNOLOGY |
MYCOLOGY |
PARASITOLOGY |
VIROLOGY |
|
ENGLISH
|
BAKTERİYOLOJİ – BÖLÜM ON
BAKTERİYEL PATOGENEZİNİN GENEL ÖZELLİKLERİ
Dr Alvin Fox
Emeritus Professor
University of South Carolina School of Medicine
Çeviren.
Prof. Dr. Mustafa Demirci
İzmir Katip Çelebi Üniversitesi,
Tıp Fakültesi, Tıbbi Mikrobiyoloji A.D.
|
|
SPANISH |
|
ALBANIAN |
|
|
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|

 |
|
Logo image © Jeffrey
Nelson, Rush University, Chicago, Illinois and
The MicrobeLibrary |
|
|
|
ANAHTAR KELİMELER,
Patojen/salgın, normal flora, enfeksiyon, enfeksiyon hastalığı, duyarlı
konak, fırsatçı enfeksiyon, nozokomiyal bulaşma, Koch postülatları, adezyon
(yapışma), penetrasyon (nüfuz etme), invazyon / yayılma, hücre dışı/ hücre
içi parazit, kapsül, ekzotoksin, endotoksin, immünopatoloji, otoimmunite.
|
Patogenez, konağın bağışıklık durumu, enfeksiyon etkeninin türü veya suşunun
doğası (hastalık oluşturma, virulans faktörleri) ve ilk maruziyette organizma
sayısına göre değişen çok faktörlü bir süreçtir.
Sınırlı sayıda bakteri türü, sağlıklı bireylerde gelişen enfeksiyon
hastalıklarının çoğunluğundan sorumludur. Aşılama, antibiyotikler ve etkili halk
sağlığı önlemlerinin başarısı nedeniyle, yakın zamana kadar olan salgınların
geçmişte olan bir şey olduğu düşünülmektedir. Antibiyotiklere dirençli
organizmaların gelişmesi nedeniyle, bu durum hızla değişmektedir.
Tüm insanlar, (deri, bağırsak ve akciğer dahil) kendi dış yüzeylerinde
yaşayan bakterilerle (normal flora) ile enfekte edilir. Biz sürekli olarak (hava,
su, toprak ve gıda dahil) bakterilere maruz kalmaktayız. Normalde, bizim sahip
olduğumuz konak savunma mekanizmaları nedeniyle bu bakterilerin çoğu zararsızdır.
Duyarlı hastalarda, bu konak savunması zayıflar ve florada bulunan bakteriler
kan dolaşımına girdiklerinde, (ameliyat, kateterizasyon veya diğer tedavi
uygulamalarından sonra) genellikle fırsatçı enfeksiyon hastalıklarına neden
olurlar. Bulaşıcı hastalıklar, hastanede başladığı zaman, hastane enfeksiyonu (nozokomiyal)
olarak adlandırılır. Normal florada, bazı yaygın bakteriler; (deride bulunan)
Staphylococcus aureus, S. epidermidis ve Propionibacterium acnes ve
bağırsaklarda Bacteroides ve Enterobacteriaceae (ikincisi çok küçük sayılarda)
bulunur.
KOCH POSTULATLARI (MODİFİYE)
1. Organizma her zaman bulaşıcı hastalığı olan insanlarda bulunmalı ancak
sağlıklı kişilerde olmamalıdır
2. Organizma bulaşıcı hastalığı olan insanlardan izole edilmeli ve saf
kültürde üretilmelidir.
3. Saf kültürde izole edilen organizma, duyarlı hayvanlara inoküle
edildiğinde hastalığı yeniden başlatmalıdır.
4. Organizma deneysel olarak enfekte hayvanlardan tekrar izole edilmelidir.
Postulatların 3. ve 4. maddeleri, insan hastalığında ajan rolü kesinliğinin
kanıtı için son derece önemlidir. Ancak, bu insan hastalığına benzer hayvan
modellerinin geliştirilmesi yeteneğine bağlıdır. Bir çok durumda bu tür modeller
yoktur.
TRANSMİSYON (BULAŞMA)
Spesifik bakteri türleri (veya türün içinde suşları), insan vücudunda belirli
bölgelere farklı yollarla bulaştıktan sonra enfeksiyon başlar. Örneğin,
bakteriler havadaki damlacıklar ile solunum yoluna, yiyecek veya suyun
tüketilmesi ile yada cinsel temas ile bulaşabilir.
|
 E. coli fimbrialarıyla (TEM x17,250) © Dennis Kunkel
Mikroskopi, Inc. izni ile kullanılır
E. coli fimbrialarıyla (TEM x17,250) © Dennis Kunkel
Mikroskopi, Inc. izni ile kullanılır |
ADEZYON (tutunma, yapışma)
Bakteriyel enfeksiyonlar genellikle konağın belirli bir epitel yüzeyine
mikrobun yapışmasıyla başlar. Aksi takdirde, bulaşan organizma
uzaklaştırılır, örneğin; peristaltizm ve dışkılamayla (bağırsaklardan),
hapşırma, öksürük ve siliyer aktiviteyle (solunum yolundan) idrar yapmakla (ürogenital
kanaldan). Yapışma non-spesifik "yapışkanlık" değildir. Bakteri hücresinin
hücre dışı bileşenleri (adhezinler) ve konak hücrenin (reseptörler) arasında
belirli etkileşimler olur. örn.; adhezin-reseptör etkileşimi.
S. pyogenes'in yüzey fimbriyası iki önemli bileşeni M proteini ve
lipoteikoik asit içerir. Fibronektin proteini epitel hücreleri ile bağlanır
ve lipoteikoik asidin yağlı asit kısımları fibronektin ile etkileşime
girerler.
Farklı yüzey özellikleri ile E. coli suşları farklı hastalıklara neden
olurlar. E. coli'nin pili yapısı araştırmaları en kapsamlı araştırmalar
arasındadır. E.coli'nin fimbriyalarının uçlarında bulunan tanımlanmış
adhezinler epitel hücrelerine bağlanmayı kolaylaştırır. Tip 1 fimbria mannoz
içeren reseptörlere bağlamanır. P fimbriyumlar glikolipit içeren galaktoza (örn.
serebrosidler) bağlanma yeteneğindedirler, epitel hücresi yüzeyinde
glikoproteinler bulunmaktadır. Başlangıçta insan eritrositleri P kan grubu
antijenlerine bağlandığı gösterildiği için "P" fimbriya olarak adlandırılır.
|
| |
PENETRASYON VE YAYILIM
Bazı bakteriyel patojenler Vibrio kolera gibi epitel yüzeylerde bulunurlar.
Diğer türler bu engellere penetre olabilirler ancak yerel kalmaları mümkündür.
Diğerleri kana veya oradan başka sistemik bölgelere geçmektedirler. Bu
genellikle bağırsak, idrar yolu ve solunum yollarında meydana gelir ve daha az
yaygınlıkta deriden olabilmektedir. Örneğin, Shigella endositik etkili olmak
için bağırsak epitel hücrelerine penetre olur; bununla birlikte Shigella
genellikle kan dolaşımına yayılmaz. Diğer bazı durumlarda, bakteriler (örneğin,
Salmonella typhi) kana epitel hücreleri içine geçmektedir. Bu durumda, invazyon
bazı durumlarda bakterinin sistemik damar içine geçmesi anlamına gelse de,
aslında invazyon bir organizmanın bir hücreye girme kabiliyetini ifade edebilir.
Borrelia bir kene ısırığı ile deri yoluyla kana bulaşır. Bazı bakteriler (örn.
hiyalüronidaz veya kollajenaz) tarafından salgılanan bazı degradatif
ekzotoksinler bağ dokusu aracılığıyla bakterilerin geçişini kolaylaştırmak için
bağ dokusu matriksini gevşetebilir.
KONAKTA YAŞAYABİLMEK
Birçok bakteriyel patojen, lizozimin veya tek başına komplemanın (alternatif
yol) veya kompleman (klasik yol) ve antikor içeren diğer vücut sıvılarının ve
plazmanın sitotoksik etkisine direnme yeteneğine sahiptir. Hücre dışı
patojenlerin yok edilmesi, büyük ölçüde opsonizasyondan (antio ve/veya kompleman
ile) sonra fagositoz sonucu fagositler içinde gerçekleşir. Hücre dışı patojenler
için fagositoz tuzağı büyük bir hayatta kalım mekanizmasıdır. Bu mekanizmadan
kaçabilmek için kapsül (birçok patojende), protein A (S. aureus) ve M proteini
(S. pyogenes) gibi yapılar fonksiyona sahiptir. Protein A, S. aureus
tarafından bir yüzey oluşturmak için salgılanan bir ürün olup, immünoglobülinin
Fc kısmına bağlanır. Bakteriler, antikorla bağlanma sonucu, C3 parçalarının
bağlanması ile sonuçlan bir süreçte klasik yoldan kompleman kaskatını aktive
ederler. Fagositoz, C3-bölgesi veya IgG'nin Fc kısmında uygun resöptörlere
opsonize bakterilerin bağlanmasından sonra ortaya çıkar. Protein A bir anti-komplemanterdir
(IgG'ye bağlanması üzerine kompleman düzeylerini tüketen kompleman kaskatı
devreye girdiği için). Bu nedenle, protein A varlığında (bağlı kompleman
aracılığıyla) C3 reseptörleri ile bakteri etkileşimi inhibe olacaktır. Serbest
protein A IgG'nin Fc kısmına bağlanır ve sonunda Fc reseptörleri aracılığıyla
fagositoz yapısal engelleme nedeniyle olmayabilir. Peptidoglikan,
lipopolisakaridler gibi, alternatif kompleman kaskatını aktifleştirebilir. S.
pyogenes de peptidoglikan yeterince açıktır ve bu komplemanı bağlayabilir. A
grubu streptokokların M proteini fimbriyasının anti fagositik bileşenidir. M
proteini, altta uzanan peptidoglikan tabakaya kopleman bağlanmasını bloklayacak
şekilde plazmadan fibrinojeni bağlar. Dolayısıyla streptokoklar non-immün serum
içinde fagosite edilemez Hücre içi patojenler (zorunlu ve fakültatif her ikisi
de) fagolizozom içinde öldürülmeden kaçabilmeldir. Bu durum, bu veziküllerden
kaçış veya parçalanması ve sonra sitoplazmada serbest yaşayabilme kabiliyeti ile
oluşabilir. Alternatif olarak, bunlar fagosomların içinde yaşayabilirler (lizozomların
fagozomlarla füzyonu inhibe edilebilir veya eğer lizozomlar ile füzyon oluşursa
organizma parçalayıcı enzimlere karşı dirençli olabilir).
|
| |
DOKU YARALANMASI
Bakteriler öncelikle farklı mekanizmalarla doku yaralanmasına neden
olabilirler:
- Ekzotoksinler
- Endotoksinler ve spesifik olmayan bağışıklık
- Spesifik humoral ve hücresel bağışıklık
Ekzotoksinler
Birçok bakteriler, bir enzimatik etkiyle, belirli hücresel yapıları
yok edebilen veya değiştirebilen, proteinler (ekzotoksinler) meydana
getirirler. Ekzotoksinlerin etkileri genellikle akut görülür ve
yeterince güçlü olduklarından ciddi etkilere (örn, ölüm) sıklıkla neden
olurlar. Bunun örneklerine; botulizm, şarbon, kolera ve difteri
verilebilir. Eğer konak akut enfeksiyondan sağ kurtulursa, sıklıkla
ekzotoksin etkisini nötralize eden konak nötralizan antikorlar (anti-toksinler)
ortaya çıkarlar. Ekzotoksin sınıfları aşağıda verilmiştir:
Bağ dokusunun hücre dışı matriksi
üzerinde etkili olan toksinler
örn; Clostridium perfringens kollajenaz, Staphylococcus aureus
hiyalüronidaz.
Bir hücreye bağlanıcı "B" ve enzimatik
olarak aktif "A" bileşeni (AB tipi toksinler) içeren toksinler
Bunlar:
a) ADP-ribozilatın aktifleşmesiyle, örn; kolera toksini, E. coli
ısıya labil toksin, Pseudomonas aeruginosa ve difteri toksinleri.
b) 28S rRNA üzerine litik etkiyle, örn; shiga ve shiga-benzeri (vero)
toksinler.
c) Kısmen belirli bölgelere etkiyle, örn; Botulinum toksini,
tetanoz toksini ve şarbon letal toksin.
Membrana hasar veren toksinler
Örn; Staphylococcus aureus delta toksin
Ekstraselüler etki yapan toksinler
Bunlar; proteazları, kolajenazları ve hiyaluronidaz'ları
içerirler. Örn; Staphylococcus aureus hiyaluronidaz üretirken,
Clostridium perfringens güçlü bir kolajenaz üretir. (Hiyalüronidaz
ve Kolajenazla) bağ doku matriksinin hasarlanması, organizmanın
daha kolay dokulara yayılmasına izin vermek için, doku liflerini
"gevşetmek" olabilir. Ayrıca, bu gruba dahil bir toksin,
epidermis içinde tabakaların ayrılmasına neden olan ve yeni
doğanlarda haşlanmış deri sendromu nedeni olan Staphylococcus
aureus exfolyatindir.
A-B toksinler
Bu tür toksinler iki bileşenden oluşur. Biri hücre yüzeyine
bağlanır ve diğeri etkisini göstereceği yere hücre zarı ya da
sitoplazma içine geçer. Bu şekilde etki gösterilen klasik
toksinler kolera ve difteri toksinleridirler.
(i) ADP-ribozile edici eksotoksinler
Difteri toksini (Corynebacterium diphtheriae tarafından
üretilir), faj toksin geni ile kodlanır. Toksin bir
polipeptit zinciri olarak sentezlenir ve hızlıca, bir
disülfid bağı ile bir arada tutulan bu polipeptid, iki
zincire (A ve B) ayrıştırılır. B hücrelere bağlanken, A
enzimatik aktiviteye sahiptir. A endositozla hücre içine
girer ve endozomdan hücre sitozolüne geçer. Difteri toksini,
ribozomlarda uzama faktörünü (EF2) ADP-ribosilatlar ve bu
nedenle protein sentezi inhibe olur. Pseudomonas ekzotoksin
A'nın, difteri toksini etkisine benzer bir şekli vardır.
Kolera toksininin çeşitli alt-birimleri bulunur ve
merkeze yerleştirilmiş bir A alt-birimi ile birlikte bir
halka oluşturabilirler. B hücre yüzeyinde gangliosidlere
bağlanır ve A'nın hücre içine penetrasyonunu sağlamak için
bir kanal oluşturur. A1, proteolitik bölünme ile meydana
getirilir ve kullanıma hazır olduktan sonra hücre zarı
regülatör kompleksi ADP-ribosilatlanır (substrat olarak NADH
kullanarak) ve sonuçta adenilat siklaz aktivasyonuna neden
olur. Adenilat siklazın aktivasyonu, siklik AMP üretiminde
bir artışa neden olur ki; bağırsak lümeninde sodyum klorür
alımında azalma ve sonucunda aktif iyon ve su
salgılanmasıyla sulu ishale neden olur. E. coli toksini
benzer bir etki şekline sahiptir.
(ii) 28S rRNA üzerinde etkili toksinler
Shiga toksinleri (kromozomaldir) şigelloz patogenezinde rol
oynarlarken, Shiga-benzeri toksinler (faj aracılıdır) esas
olarak enterohemorraghic E. coli tarafından
üretilmektedirler. Onlar ortak etki şekline sahiptirler. A
alt-biriminin A parçası, tek bir adenosin kalıntı üzerinde
N-glikosidaz aktivitesine sahip ribozoma gider. Örneğin baz
ve riboz arasındaki bağ parçalanır.
İshal aktif iyon/su salgılanması sonucunda olmaz, zayıf
su emme sonucunda olur bu da epitel hücrelerinin hücre
ölümüne yol açan protein sentezinin inhibisyonu sonucundur.
(iii) kısmen özellikli alanlara etkili toksinler
Botulinum nörotoksinleri, tetanospasmin ve B. anthracis
letal toksin AB tip ekzotoksinler olarak görünürler.
Botulinum toksini nöromüsküler kavşakta asetilkolinin
salınımının inhibisyonuna neden olarak etki eder. Tetanoz
toksini nöromusküler kavşaklarda görev yapar ve sinapslara
aksonlarla taşınır. Daha sonra inhibitör nöronları inaktive
ederek etki ederler. Tetanoz ve botulizm ekzotoksinlerinin B
bileşenleri var görünse de onların A alt birimlerinin etki
şekli bilinmemektedir. B. anthracis letal toksini B bileşeni
koruyucu bir antijendir. İlginç olarak, bu aynı zamanda ödem
toksin B alt birimi olarak hizmet vermektedir.
Membrana hasar verici toksinler
Bu toksinler, enzimatik olarak membran fosfolipid (veya proteini)
bileşenleri sindirir veya deterjan olarak etki ederler. Heriki
durumda da hücre zarında zımbalanmış gibi delikler açılmıştır ve
sitoplazmik içerikleri süzülebilir. C. perfringens fosfolipazı
("toksin"), membran hasarı oluşturan toksinlere bir örnektir. Bu
kan damarlarını tahrip eder ve inflamatuar hücrelerin akışını
durdurur. Bu, aynı zamanda, zorunlu anaerob üremesinde önemli
olan bir anaerobik ortam oluşturulmasına yardımcı olur. S.
aureus delta toksini, hücre zarları içine yerleştirilen ve bir
deterjan benzeri etkiye sahip olduğuna inanılan, büyük
hidrofobik bir proteindir.
|
| |
Endotoksinler
Antibiyotik çağın avantajlarına karşın, her yıl yaklaşık 200.000 hastada
Gram negatif sepsis gelişir ve bunların yaklaşık % 25-40'ı septik şok ve
ölümle sonuçlanır. Septik şokta (sıvıların dokularda havuzlanması nedeniyle
) hipotansiyon, yaygın damar içi pıhtılaşma ve ateş gelişir sonuçta sıklıkla
yaygın sistem yetmezlikleri nedeniyle ölümcüldür. Aynı zamanda beyin gibi
hassas dokularda etkili olan oksijenlenme eksikliği gelişmektedir.
Hastalarda lipid A veya peptidoglikanın toksik aktivitesini ters çevirecek
etkili bir tedavi henüz yoktur.
Endotoksinler, bakteri hücre zarfının toksik bileşenleridir. Klasik ve en
güçlü endotoksin lipopolisakkariddir. Ancak, peptidoglikan birçok endotoksin
benzer özelliklere sahiptir. Bazı peptidoglikanlar zayıfça biyolojik olarak
parçalanabilirler ve kronik yanı sıra akut doku yaralanmasına da neden
olabilirler. Endotoksinler inflamasyonu "non-spesifik" kışkırtırlar. Örneğin,
bağışıklık sistemi hücreleri heryerde (interlökin-1 ve tümör nekroz faktörü
de dahil olmak üzere) sitokinleri serbest bırakmaları için uyarılırlar.
Endotoksinler da alternatif kompleman yolunu etkinleştir. Etkilenmiş
dokulara polimorfonükleer hücrelerin çekimi bu sitokinlerin üretimi
sonucunda artar. PG ve LPS ve bazı diğer hücre duvarı bileşenleri (örneğin,
pnömokok kökenli teikoik asit) da alternatif kompleman kaskatının
aktivatörleridir. Böylece birçok bakteri komplemanı bağlar ve fagositlerce
kendi alımını teşvik eder ve antikor yokluğunda öldürülebilir. Bazı
kompleman yan ürünleri ayrıca, nötrofillerin kemoatraktanlarıdırlar (kimyasal
çekicileridir). Endotoksinler ayrıca potent B hücresi mitojeni, poliklonal B
hücresi aktivatörü ve adjuvanıdırlar (heriki antikor ve hücre aracılı
bağışıklık için); bu, mikroplar akut olarak ortadan kaldırılmazsa, onlarla
mücadelede uygun bir kronik bağışıklık cevabının gelişmesinde önemli bir rol
oynamaktadır.
Akut faz süresince bir "birincil" enfeksiyonu "non-antijen spesifik" bir
şekilde immun sistemin ortadan kaldırması son derece önemli olacaktır.
Organizma devam ederse (veya daha sonraki zamanda bir reinfeksiyon olursa),
spesifik bir bağışıklık organizmaların gelişimini yavaşlatma veya
enfeksiyonun ortadan kaldırılmasında daha büyük değere sahip olacaktır. Bu
durum, tüberküloz, cüzam, Lyme hastalığı ve frengi gibi kronik
enfeksiyonlarda önemlidir.
İmmunopatoloji
Enfekte doku genellikle masum bir seyirci olarak immünopatoloji
sonuçlarından etkilenir. Bu akut veya kronik enfeksiyonlarda da oluşabilir.
Endotoksinler tarafından sitokin üretimi ve kompleman aktivasyon uyarılması
süresince, bir immun yanıtının yokluğunda da doku yaralanması oluşabilir.
İnatçı canlı mikroplardan sürekli serbest antijenler oluşturulduğunda,
ardından hümoral antikorlar ve hücre aracılı bağışıklığın uyarılması ve
kronik immunopatolojiler ile sonuçlanır. Bazı kötü parçalanabilir antijenler
olduğunda (örneğin, pnömokok kökenli polisakarid ve A grubu streptokok hücre
duvarları), hatta canlı ajanların sürekliliğinin yokluğunda,
immunopatalojiyi devam edebilir. Bazı diğer bakteriyel antijenler konakçı
doku antijenleri ile otoimmünite oluşumuna yol açacak şekilde çapraz
reaksiyona girebilirler (örneğin, S. pyogenes M proteini memeli miyozini ile
çapraz reaksiyona neden olur). Böylece immünopatoloji, enfeksiyonun ve
mikrobiyal antijenlerin ortadan kaldırılmasından sonra bile devam edebilir.
Enfeksiyona karşı bağışıklık
sistemi - örnekler
- Hücre dışı parazitler. Antikorlar, organizmanın parçalanması ve/veya
bunların fagositler tarafından hızla öldürülmesini sağlayan
işaretlenmesiyle opsonizasyonuna neden olurlar.
- Hücre içi parazitler öncelikle hücre aracılı immünite tarafından
öldürülür.
- Ekzotoksinler antitoksinler tarafından nötralize edilebilir.
Antitoksinler toksoid aşılar (toksoidler antijenik ama zararlı değildir)
kullanılarak elde edilebilir. Bu difteri aşı örneğinde ortaya çıkar.
- Bazı organizmalar IgA proteazlar üretirler. Bu dış yüzeyler üzerinde
hayatta kalmaya yardımcı olur (H. influenzae, S. pneumoniae, N.
gonorrhoeae ve N. meningitidis de dahil olmak üzere).
|
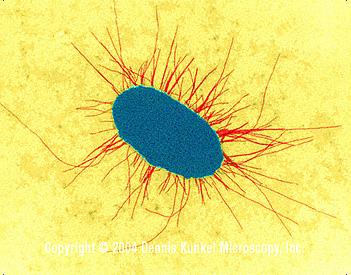 E. coli fimbrialarıyla (TEM x17,250) © Dennis Kunkel
Mikroskopi, Inc. izni ile kullanılır
E. coli fimbrialarıyla (TEM x17,250) © Dennis Kunkel
Mikroskopi, Inc. izni ile kullanılır