| xx | x | ||||
|
|
 |
||||
| INFECTIOUS DISEASE | BACTERIOLOGY | IMMUNOLOGY | MYCOLOGY | PARASITOLOGY | VIROLOGY |
|
|||||
| ALBANIAN | |||||
|
Let us know what you think FEEDBACK |
|||||
| SEARCH | |||||
|
|
|||||
|
Logo image © Jeffrey Nelson, Rush University, Chicago, Illinois and The MicrobeLibrary |
|||||
|
|
|||||
|
Key
Words |
Pathogenesis is a multi-factorial process which depends on the immune status of the host, the nature of the species or strain (virulence factors) and the number of organisms in the initial exposure. A limited number of bacterial species are responsible for the majority of infectious diseases in healthy individuals. Due to the success of vaccination, antibiotics, and effective public health measures, until recently, epidemics were felt to be a thing of the past. Due to the development of antibiotic resistant organisms, this situation is changing rapidly. All humans are infected with bacteria (the normal flora) living on their external surfaces (including the skin, gut and lungs). We are constantly also exposed to bacteria (including air, water, soil and food). Normally due to our host defenses most of these bacteria are harmless. In compromised patients, whose defenses are weakened, these bacteria often cause opportunistic infectious diseases when entering the bloodstream (after surgery, catheterization or other treatment modalities). When initiated in the hospital, these infectious diseases are referred to as nosocomial. Some common bacteria found in the normal flora include Staphylococcus aureus, S. epidermidis and Propionibacterium acnes (found on the skin) and Bacteroides and Enterobacteriaceae found in the intestine (the latter in much smaller numbers).
Koch's postulates (modified) 1. The organism must always be found in humans with the infectious disease but not found in healthy ones. 2. The organism must be isolated from humans with the infectious disease and grown in pure culture. 3. The organism isolated in pure culture must initiate disease when re-inoculated into susceptible animals. 4. The organism should be re-isolated from the experimentally infected animals. Postulates 3. and 4. are extremely important in definite proof of the role of agent in human disease. However, this depends on the ability to develop animal models that resemble the human disease. In many cases such models do not exist. Transmission Specific bacterial species (or strains within a
species) initiate infection after being transmitted by different routes to
specific sites in the human body. For example, bacteria are transmitted in
airborne droplets to the respiratory tract, by ingestion of food or water or by
sexual contact. Adhesion Bacterial infections are usually initiated by adherence of the microbe to a specific epithelial surface of the host. Otherwise the organism is removed e.g. by peristalsis and defecation (from the gut) ciliary action, coughing and sneezing (from the respiratory tract) or urination (from the urogenital tract). Adhesion is not non-specific "stickiness". Specific interactions between external constituents on the bacterial cell (adhesins) and on the host cell (receptors) occur i.e. an adhesin-receptor interaction. S. pyogenes has surface fimbriae which contain two major components the M protein and lipoteichoic acid. The protein fibronectin binds to epithelial cells and fatty acid moieties of lipoteichoic acid in turn interact with fibronectin. |
||||
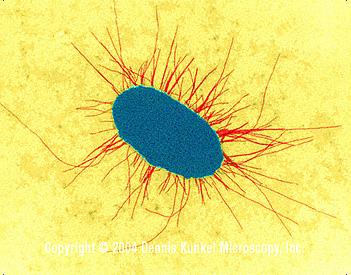 E. coli with fimbriae (TEM x17,250)
©
Dennis Kunkel Microscopy, Inc.
Used with permission
E. coli with fimbriae (TEM x17,250)
©
Dennis Kunkel Microscopy, Inc.
Used with permission |
Strains of E. coli with different surface characteristics cause distinct diseases. Among the most thoroughly studied pili are those of uropathogenic E. coli. Certain adhesins present on the tips of fimbriae of E. coli facilitate binding to epithelial cells. Type 1 fimbriae bind to mannose containing receptors. Whilst P fimbriae allow binding to galactose containing glycolipids (e.g. cerebrosides) and glycoproteins present on epithelial cells. They are referred to as "P" fimbriae since they were originally shown to bind to P blood group antigens on human erythrocytes. | ||||
|
Penetration and spread Some bacterial pathogens reside on epithelial surfaces e.g. Vibrio cholerae. Other species are able to penetrate these barriers but remain locally. Others pass into the bloodstream or from there onto other systemic sites. This often occurs in the intestine, urinary tract and respiratory tract, and much less commonly through the skin. For example, Shigella penetrates by activating epithelial cells of the intestine to become endocytic; the Shigella do not usually spread into the bloodstream. In other cases, bacteria (e.g. Salmonella typhi) pass through epithelial cells into the bloodstream. Thus, invasion can refer to the ability of an organism to enter a cell, although in some instances it can mean further passage into the systemic vasculature. Borrelia burgdorferi is transmitted into the bloodstream through the skin by a tick bite. Certain degradative exotoxins secreted by some bacteria (e.g. hyaluronidase or collagenase) can loosen the connective tissue matrix increasing the ease of passage of bacteria through these sites. Survival in the host Many bacterial pathogens are able to resist the cytotoxic action of plasma and other body fluids involving antibody and complement (classical pathway) or complement alone (alternate pathway) or lysozyme. Killing of extracellular pathogens largely occurs within phagocytes after opsonization (by antibody and/or complement) and phagocytosis. Circumvention of phagocytosis by extracellular pathogens is thus a major survival mechanism. Capsules (many pathogens), protein A (S. aureus) and M protein (S. pyogenes) function in this regard. Protein A is a surface constituent of S. aureus as well as a secreted product and binds to the Fc portion of immunoglobulins. Bacteria, on binding antibody, activate the classical complement cascade which results in the attachment of fragments of C3. Phagocytosis occurs after binding of the opsonized bacteria to receptors for the Fc portion of IgG or C3 regions. Protein A is anti-complementary (since, on binding to IgG, the complement cascade is activated, depleting complement levels). Thus in the presence of protein A, interaction of bacteria (via bound complement) with C3 receptors will be inhibited. Free protein A binds to the Fc portion of IgG, thus phagocytosis via Fc receptors may not occur because of steric hindrance. Peptidoglycan, like lipopolysaccharide, can activate the alternate complement cascade. In S. pyogenes peptidoglycan is sufficiently exposed that it is able to bind complement. The M protein of group A streptococci is the anti-phagocytic component of the fimbriae. M protein binds fibrinogen from plasma which blocks complement binding to the underlying peptidoglycan layer. Thus streptococci in non-immune serum are not phagocytosed. Intracellular pathogens (both obligate and facultative) must be able to avoid being killed within phagolysozomes. This can occur from by-passing or lysing these vesicles and then residing free in the cytoplasm. Alternatively, they can survive in phagosomes (fusion of phagosomes with lysosomes may be inhibited or the organism may be resistant to degradative enzymes if fusion with lysosomes occurs). Tissue injury Bacteria cause tissue injury primarily by several distinct mechanisms involving:
|
|||||
|
|||||
|
|||||