|
x |
x |
|
 |
 |
|
BỆNH NHIỄM TRÙNG |
VI KHUẨN HỌC |
MIỄN DỊCH HỌC |
NẤM HỌC |
KÝ SINH TRÙNG HỌC |
VIRÚT HỌC |
|
ENGLISH |
MIỄN DỊCH HỌC – CHƯƠNG SÁU
GEN HỌC CỦA KHÁNG THỂ
Gene Mayer, Ph.D
Emertius Professor of Pathology, Microbiology and Immunology
University of South Carolina
Biên dịch: Nguyễn Văn Đô, MD., PhD.,
Bộ môn Sinh lý bệnh-Miễn dịch,
Trường Đại học Y Hà Nội,
Hà Nội, Việt Nam
|
|
TURKISH |
|
FRANCAIS |
|
PORTUGUES |
|
SHQIP |
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|

 |
|
Logo image
© Jeffrey Nelson, Rush University, Chicago, Illinois and
The MicrobeLibrary |
|
|
|
|
|
MỤC TIÊU GIẢNG DẠY
Mô tả cấu trúc và biểu lộ của gia đình gen kháng thể
Giải thích nguồn gốc sự đa dạng của kháng thể
|
LỊCH SỬ
Dữ liệu về trình tự acid amin cho thấy rằng một vùng C duy nhất
có thể kết hợp với nhiều vùng V khác nhau. Ngoài ra, người ta cho rằng một
idiotyp duy nhất có thể được liên kết với các vùng C khác nhau (ví dụ: IgM và
IgG). Để giải thích những cở sở dữ liệu này có ý kiến cho rằng có lẽ hai vùng
của phân tử kháng thể đã được mã hoá bởi các gen riêng biệt và các gen vùng V và
C được nối với nhau bằng cách nào đó trước khi một phân tử kháng thể được hình
thành (tức là có hai gen mã hóa cho một polypeptid). Đây là một khái niệm mang
tính cách mạng nhưng với sự ra đời của công nghệ DNA tái tổ hợp, nó đã cho thấy
khái niệm đó là đúng. Các chuỗi nặng và chuỗi nhẹ của kháng thể được mã hoá bởi
các gia đình ba gen khác nhau, mỗi một gen riêng biệt trên một nhiễm sắc thể
riêng biệt - một cho chuỗi nặng và một cho mỗi loại chuỗi nhẹ. Mỗi một gia đình
các gen này có một số gen cho một vài vùng V và một hoặc nhiều gen cho vùng C.
Tuy nhiên, các gen vùng V và C không phải nằm liền kề với nhau.and la
|
TỪ KHÓA
Gen V
Gen C
Vùng J
Vùng D
Chất dẫn đầu
Chất tăng cường
Promotor
Sự đa dạng kháng thể
Thuyết dòng mầm
Thuyết đột biến tế bào soma
Chèn vùng N
Đa dạng liên kết
Phối hợp kết hợp
Đa đặc hiệu
Chọn lọc dòng
|
GIA ĐÌNH GEN CHUỖI NHẸ
Tổ chức gen ở dòng mầm
Việc tổ chức các gen chuỗi nhẹ Kappa và
Lambda trong dòng mầm của các tế bào chưa biệt hoá được mô tả trong Hình 1.
-
Chuỗi nhẹ
Lambda
Gia đình gen lambda gồm có 4 gen vùng C, mỗi gen cho một dưới nhóm
của chuỗi lamda, và khoảng 30 gen vùng V. Mỗi gen trong số các gen
vùng V có hai exon, một (L) mã hóa cho vùng chính và gen khác (V) mã
hóa cho hầu hết vùng thay đổi. Theo chiều ngược mỗi gen của các gen
vùng C có thêm một exon được gọi là J (joining:chỗ nối). Các exon L,
V, J và C được ngăn cách bởi các intron (các trình tự không mã hóa
xen vào giữa).
-
Chuỗi nhẹ
Kappa
Gia đình gen chuỗi nhẹ Kappa chỉ có một gen vùng C duy nhất, vì chỉ
có một loại chuỗi nhẹ Kappa. Có rất nhiều gen vùng V (khoảng 250)
mỗi gen có exon chính và một exon V. Trong gia đình gen kappa có một
số exon J nằm giữa gen V và C. Tất cả các exon được phân cách bởi
các intron.
|
 Hình 1
Hình 1
Tổ chức của các gen chuỗi nhẹ kappa và lamda ở dòng mầm
hoặc tế bào chưa biệt hóa
|
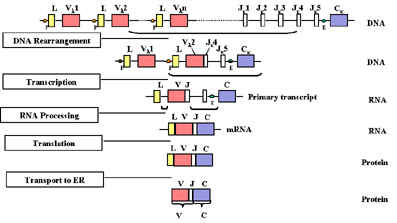
Sắp xếp lại gen và biểu lộ gen
Khi một tế bào biệt hóa thành tế bào lympho B trưởng
thành, nó sẽ tổng hợp ra một chuỗi nhẹ, có sự sắp xếp lại các gen (exon)
và gen này bắt đầu được biểu hiện được mô tả trong Hình 2.
|
 Hình 2
Hình 2
Khi một tế bào biệt hóa thành tế bào B chín nó sẽ tổng
hợp ra chuỗi nhẹ và có sự tái sắp xếp một số gen (exon)
|
Khi một tế bào trở thành tế bào B, nó sẽ tạo
ra một chuỗi nhẹ và có sự sắp xếp lại các gen ở mức độ DNA sao cho một trong
những gen V được đưa đến bên cạnh một gen của các vùng J. Điều đó xuất hiện
nhờ sự tái tổ hợp bằng cách loại bỏ intron giữa các vùng V và J. Việc lựa
chọn gen V nào là hoàn toàn ngẫu nhiên; có một số ưu tiên cho việc sử dụng
các gen V gần nhất với vùng J. Tuy nhiên, cùng với thời gian tất cả các gen
V có thể được sử dụng và tất cả các phối hợp của các gen V và các vùng J
được thực hiện.
Kết quả của sự sắp xếp lại DNA này là gen
trở nên hoạt hóa phiên mã vì promoter (P) liên quan đến gen V được đưa
gần với chất kích hoạt (E), nằm trong intron giữa các vùng J và C. Khi
phiên mã bắt đầu từ promoter, tiền mRNA được tổng hợp có chứa các trình
tự nucleotid từ các vùng L, V J và C cũng như trình tự của intron giữa L
và V và giữa J và C (Hình 2). Tiền mRNA này được xử lý (spliced) trong
nhân tế bào và các intron còn lại được loại bỏ. Cuối cùng mRNA sẽ có các
exon L, V J và C tiếp giáp nhau.
mRNA được dịch mã ở bào tương và chất dẫn
đầu được gỡ bỏ khi protein được vận chuyển vào lumen của lưới nội chất.
Chuỗi nhẹ được lắp ráp với một chuỗi nặng trong lưới nội chất và kháng
thể được tiết ra thông qua con đường tiết protein bình thường. Vùng V
của chuỗi nhẹ trưởng thành được mã hoá bởi trình tự nucleotid trong gen
V và vùng J và vùng C bởi các trình tự trong gen C.
|
 Hình 3 Ngoài một số exon J, gia đình gen chuỗi nặng
cũng có thêm một số exon được gọi là các exon D (đa dạng). Tất cả các
exon được ngăn cách bởi intron
Hình 3 Ngoài một số exon J, gia đình gen chuỗi nặng
cũng có thêm một số exon được gọi là các exon D (đa dạng). Tất cả các
exon được ngăn cách bởi intron
|
GIA ĐÌNH GEN CHUỖI NẶNG
Tổ chức gen dòng mầm
Sự tổ chức các gen chuỗi nặng được mô tả trong Hình 3.
Trong gia đình gen chuỗi nặng có
nhiều gen C, một gen cho mỗi lớp và dưới lớp của kháng thể. Trên
thực tế mỗi một gen của các gen vùng C bao gồm một số exon, một exon
cho mỗi domain và một exon khác cho vùng bản lề. Trong gia đình gen
chuỗi nặng có nhiều gen vùng V, mỗi gen được hình thành bởi một gen
dẫn đầu và exon V. Ngoài một số exon J, gia đình gen chuỗi nặng cũng
có chứa một vài exon được gọi là exon D (diversity:đa dạng). Tất cả
các exon được phân cách bởi các intron như mô tả trong Hình 3.
|
| |
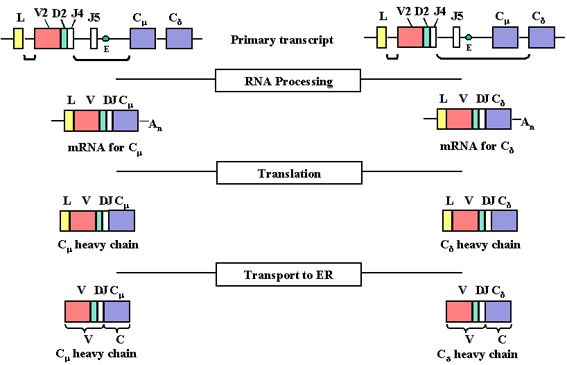
Sự tái sắp xếp và biểu lộ gen
Khi một tế bào biệt hóa thành một tế bào B trưởng
thành sẽ sản xuất một chuỗi nặng, có một sự sắp xếp lại các đoạn gen
khác nhau (exon) và gen này bắt đầu được biểu lộ như mô tả trong
hình 4 và 5.
|
 Hình 4
Hình 4
Khi bắt đầu phiên mã từ một promoter, tiền mRNA được tạo
ra và nó chứa các trình tự nucleotid từ các vùng L, V, D, J, Cμ và Cδ
cũng như trình tự do các intron giữa L và V, giữa J và Cμ, và giữa Cμ và
Cδ
 Hình 5
Hình 5
Tiền mRNA được xử lý (cắt và ghép) ở nhân tế bào và các
intron còn lại được loại bỏ, bao gồm cả những intron nằm giữa các exon
của các gen C
|
Khi một tế bào trở thành một tế bào B sản
xuất một chuỗi nặng, có hai sự sắp xếp lại ở mức độ ADN. Trước tiên, một
trong những vùng D được đưa đến bên cạnh một trong những vùng J và sau
đó một trong những gen V được đưa đến bên cạnh khu vực DJ đã được sắp
xếp lại. Điều này xảy ra bởi hai sự kiện tái tổ hợp trong đó loại bỏ các
intron giữa các vùng V, D và J. Cùng với các chuỗi nhẹ, sự lựa chọn của
gen V chuỗi nặng không phải là hoàn toàn ngẫu nhiên nhưng cuối cùng tất
cả các gen V có thể được sử dụng.
Kết quả của sự tái sắp xếp lại DNA này là
gen sẽ trở nên hoạt hóa phiên mã nhờ một promoter (P) (liên kết với các
gen V) được đưa lại gần một chất tăng cường (E), nó nằm trong intron
giữa vùng J và Cmu . Khi phiên mã được bắt đầu từ một promoter, tiền
mRNA được tổng hợp có trình tự của các vùng L, V, D, J Cmu và Cdelta
cũng như trình tự của các intron giữa L và V, giữa J và Cmu, và giữa Cmu
và Cdelta (Hình 4).
Tiền mRNA được xử lý (cắt ghép) trong
nhân tế bào và các intron còn lại, bao gồm cả intron giữa các exon trong
gen C đều bị loại bỏ (xem Hình 5). Tiền mRNA có thể được xử lý theo hai
cách, một là mang VDJ đến bên cạnh gen Cmu và hai là mang VDJ lại cạnh
gen Cdelta. Các mRNA cuối cùng có các exon liền kề L, V, D, J và Cmu
hoặc Cdelta và sẽ mã hóa cho một chuỗi mu và một chuỗi delta một cách
tương ứng.
Các mRNA được dịch mã trong bào tương và chất dẫn đầu được cắt bỏ khi
protein được vận chuyển vào trong lumen của mạng lưới nội chất. Chuỗi
nặng được lắp ráp với một chuỗi nhẹ trong mạng lưới nội sinh chất và
kháng thể được tiết ra qua con đường tiết protein thông thường. Khu vực
V của chuỗi nặng trưởng thành được mã hoá theo trình tự trong gen V,
vùng D và vùng J, và vùng C bởi trình tự trong gen C.irs
|
 Hình 6
Hình 6
Bên cạnh các exon V, J và D có một chuỗi duy nhất được
gọi là trình tự tín hiệu tái tổ hợp (RSS), nó có chức năng tái tổ hợp.
Mỗi RSS bao gồm một nonamer và một heptamer bảo tồn được phân cách bởi
12 hoặc 23 đôi bazơ
|
CƠ CHẾ TÁI SẮP XẾP DNA
Nằm bên cạnh các exon V, J và D, có một trình
tự gen độc nhất được gọi là trình tự tín hiệu tái tổ hợp (RSS), có chức năng
tái tổ hợp. Mỗi RSS có một nonamer và một heptamer bảo tồn được phân cách
bởi 12bp hoặc 23bp như minh họa trong Hình 6. Các khoảng 12bp và 23bp tương
ứng với một hoặc hai chu kỳ của chuỗi xoắn DNA.
Tái tổ hợp chỉ xảy ra giữa tín hiệu lượt 1 và
tín hiệu lượt 2. Trong trường hợp của chuỗi nhẹ λ có 1 lượt tín hiệu ngược
dòng của exon J và tín hiệu lượt 2 xuôi chiều của Vlambda. Trong trường hợp
của các chuỗi nhẹ κ có 1 lượt tín hiệu xuôi chiều của gen Vkappa và một tín
hiệu ngược chiều lượt 2 của các exon J. Trong trường hợp của các chuỗi nặng
có các tín hiệu lượt 1 tác động trên mỗi bên của các exon D và một tín hiệu
xuôi chiều lượt 2 của gen V và một tín hiệu ngược chiều lượt 2 của các exon
J. Vì vậy, điều đó đảm bảo rằng các sự kiện tái tổ hợp chính xác sẽ xảy ra.
Sự kiện tái tổ hợp dẫn đến việc loại bỏ các
intron giữa V và J trong trường hợp của các chuỗi nhẹ hoặc giữa các V, D, và
J trong trường hợp của các chuỗi nặng. Sự kiện tái tổ hợp được thực hiện bởi
hai protein, Rag-1 và Rag-2. Những đột biến gen của các protein này sinh ra
một bệnh suy giảm miễn dịch phối hơp nặng (cả các tế bào T và B đều suy giảm),
vì các protein này và các RSS có liên quan đến hình thành thụ thể đối với
kháng nguyên của cả tế bào B và T.
|
 Hình 7
Hình 7
Trình tự biểu lộ gen Ig - Chuỗi nặng
 Hình 8
Hình 8
Trình tự biểu lộ gen Ig - Chuỗi nhẹ
|
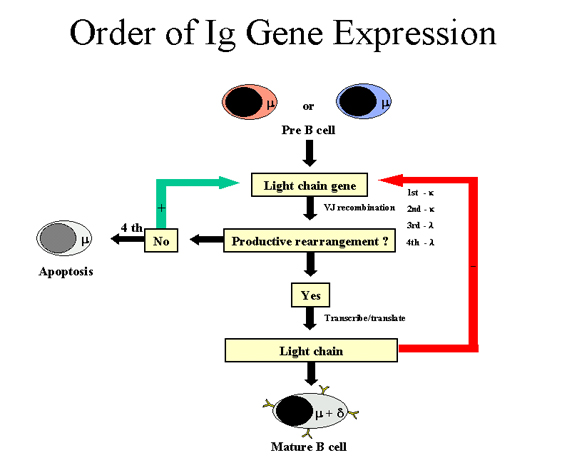
TRÌNH TỰ BIỂU LỘ GEN TRONG GIA ĐÌNH GEN GLOBULIN MIỄN
DỊCH Một tế bào B chỉ sản xuất một loại chuỗi
nhẹ và một lớp chuỗi nặng. (Lưu ý: Có một ngoại lệ là một tế bào B trưởng
thành có thể sản xuất cả hai chuỗi nặng μ và δ nhưng tính đặc hiệu của kháng
thể là tương tự vì các vùng tương tự VDJ được phát hiện ở các chuỗi μ và δ).
Vì bất kỳ tế bào B nào có cả hai nhiễm sắc thể của mẹ và cha mà mã cho các
gen kháng thể phải có một số trật tự, trong đó một tế bào biểu lộ gen kháng
thể của nó phải đảm bảo rằng chỉ có một loại chuỗi nhẹ và một lớp chuỗi nặng
được sản xuất.
Thứ tự các gen globulin miễn dịch được biểu hiện trong một
tế bào B được mô tả trong Hình 7 và 8.
Chuỗi nặng (Hình 7)
Trước hết một tế bào sẽ sắp xếp lại một trong những gen
chuỗi nặng của nó; trong một số tế bào nhiễm sắc thể của người mẹ được
chọn và ở những tế bào khác khác nhiễm sắc thể người cha được chọn. Nếu
sự tái sắp xếp thành công thì sẽ tạo ra một chuỗi nặng, sau đó sẽ không
có sự tái sắp xếp nào nữa ở các gene chuỗi nặng. Mặt khác, nếu nỗ lực
đầu tiên để sắp xếp chuỗi gen nặng không thành công (tức là không có
chuỗi nặng được sinh ra), sau đó tế bào sẽ cố gắng sắp xếp gen chuỗi
nặng trên nhiễm sắc thể khác của nó. Nếu tế bào không thành công việc
sắp xếp lại các gen chuỗi nặng lần thứ hai, đó là định mệnh sẽ được loại
bỏ.
|
| |
Chuỗi nhẹ Kappa (Hình 8)
Khi một tế bào tái sắp xếp thành công một gen
chuỗi nặng, nó bắt đầu tái sắp xếp một trong những gen chuỗi nhẹ
kappa. Nó là một sự kiện ngẫu nhiên, chuỗi nhẹ kappa từ mẹ hoặc từ
cha sẽ được chọn. Nếu sự tái sắp xếp không thành công (tức là nó
không tạo ra một chuỗi nhẹ kappa có chức năng), sau đó nó cố gắng
tái sắp xếp các gen kappa trên nhiễm sắc thể khác. Nếu tế bào tái
sắp xếp thành công một gen chuỗi nhẹ kappa, nó sẽ là một tế bào B
sản xuất một kháng thể với một chuỗi nhẹ kappa.
|
| |
Chuỗi nhẹ Lambda (Hình 8)
Nếu một tế bào tái xếp cả hai gen chuỗi nhẹ kappa
của nó không thành công, nó sẽ cố gắng để thực hiện một chuỗi nhẹ
lambda. Nó là một sự kiện ngẫu nhiên, gen chuỗi nhẹ lambda từ người
mẹ hoặc từ người cha được chọn. Nếu tái sắp xếp không thành công (tức
là nó không tạo ra một chuỗi nhẹ lambda có chức năng) thì nó cố gắng
tái sắp xếp các gen lambda trên nhiễm sắc thể khác. Nếu tế bào tái
sắp xếp thành công một gen chuỗi nhẹ lambda, nó sẽ là một tế bào B
sản xuất một kháng thể với một chuỗi nhẹ lambda.
Trình tự tái sắp xếp các gen trong các gia đình
gen kháng thể giải thích:
- Tại sao một tế bào B riêng lẻ chỉ có thể sản
xuất một loại kháng thể với một loại chuỗi nặng và một loại
chuỗi nhẹ.
- Tại sao một tế bào B riêng lẻ chỉ có thể sản
xuất các kháng thể có tính đặc hiệu duy nhất.
- Tại sao có loại trừ allel trong kháng thể
allotyp ở mức độ phân tử kháng thể riêng rẽ, nhưng biểu hiện
đồng trội của allotyp trong cơ thể nói chung.
|
 Hình 9 Nguồn gốc các khái niệm hiện nay về sự đa
dạng của kháng thể
Hình 9 Nguồn gốc các khái niệm hiện nay về sự đa
dạng của kháng thể |
NGUỒN GỐC SỰ ĐA DẠNG CỦA KHÁNG
THỂ
Cơ sở
Sự đa dạng của kháng thể được cho là
tổng số tất cả sự đặc hiệu của kháng thể mà một cơ thể tạo ra. Người
ta ước tính rằng cơ thể chúng ta có thể sản xuất 107-108 phân tử
kháng thể khác nhau. Một trong những câu hỏi quan trọng trong miễn
dịch học là làm thế nào mà cơ thể chúng ta có thể sản xuất rất nhiều
phân tử kháng thể khác nhau đó. Các giả thuyết giải thích về nguồn
gốc của sự đa dạng kháng thể được xếp vào hai loại chính.
Thuyết dòng
mầm
Lý thuyết này cho rằng cơ thể
chúng tai có một gen vùng V khác nhau cho mỗi kháng thể có thể
được sản xuất.
Thuyết đột
biến tế bào thân (soma)
Thuyết này cho rằng cơ thể chúng
ta chỉ có một hoặc một vài gen vùng V và sự đa dạng được tạo ra
bởi những đột biến soma xảy ra trong những gen này.
Khái niệm hiện tại
Suy nghĩ hiện tại của chúng ta là cả hai thuyết dòng
mầm và thuyết đột biến soma đều có lý. Người ta cho rằng tính đa
dạng kháng thể được tạo ra bởi các cơ chế sau đây.
1. Một số lượng lớn các gen V
Hiện có:
a) 30 gen V lambda
b) 300 gen V kappa
c) 1000 gen V chuỗi nặng
2. Liên hợp V-J và V-D-J
Các vùng nơi mà các gen chuỗi nhẹ V và vùng J hoặc gen V chuỗi
nặng và vùng D và J kliên kết với nhau là vùng siêu biến thứ ba.
Khi mà ngẫu nhiên V, J hay D liên kết với nhau, có rất nhiều
tính đa dạng có thể được tạo ra bởi liên kết V-J và V-D-J.
3. Phối hợp đa dạng (không chỉ
trong tái tổ hợp V-J và V-D và D-J) - (Hình 9)
Tái tổ hợp giữa V-J và V-D-J
không phải lúc nào cũng hoàn thiện và sự đa dạng có thể phát
sinh do lỗi xảy ra trong trường hợp tái tổ hợp đó sẽ đem vùng V
đến bên cạnh các vùng J hoặc D, hoặc vùng D sát vùng J. Người ta
ước tính rằng những sai sót có thể gấp ba lần sự đa dạng được
tạo ra bởi liên hợp V-J và V-D-J. Sự đa dạng được tạo ra bởi cơ
chế này đang xảy ra ở vùng siêu biến thứ ba và do đó ảnh hưởng
trực tiếp đến các vị trí kết hợp của kháng thể.
|
| |
4. Chèn vùng N
Có một đoạn nucleotid chèn vào nơi tiếp
giáp giữa các mảnh D và J được xúc tác bởi enzyme terminal transferase.
Terminal transferase xúc tác sự trùng hợp ngẫu nhiên của các nucleotide
trong DNA mà không cần một bản làm khuôn mẫu. Điều này dẫn đến sự đa
dạng hơn nữa trong khu vực siêu biến thứ ba.
5. Đột biến tế bào soma
Có bằng chứng cho rằng đột biến soma đang
xảy ra trong gen V, đặc biệt là ở nơi mà mã hóa cho các vùng siêu biến
thứ hai. Vì vậy, đột biến soma có thể góp phần vào sự đa dạng kháng thể
với mức độ nào đó.
6. Kết hợp tổ hợp
Bất kể tế bào B nào cũng có khả năng sản
xuất bất kỳ một trong những chuỗi nặng và chuỗi nhẹ. Như vậy, sự kết hợp
khác nhau của chuỗi nặng và nhẹ trong một tế bào B sẽ bổ sung thêm sự đa
dạng hơn nữa.
7. Đa đặc hiệu
Do phản ứng chéo giữa các quyết định
kháng nguyên của cấu trúc tương tự, một kháng thể thường có thể phản ứng
với hơn một quyết định KN. Điều này được gọi là đa đặc hiệu. Đa đặc hiệu
cũng góp phần vào sự đa dạng của kháng thể.
Ví dụ về cơ chế làm thế nào để có thể tạo ra sự đa
dạng lớn được minh họa dưới đây:
|
| |
|
|
Thụ thể tế bào B (kháng thể) |
|
Chuỗi nặng |
Kappa |
|
Các mảnh gen V |
1000 |
300 |
|
Các mảnh gen D |
15 |
- |
|
Các mảnh gen J |
4 |
4 |
|
Chèn vùng N |
++ |
- |
|
Đa dạng nơi kết nối |
+++ |
+ |
|
Đột biến soma |
+ |
+ |
|
Kết hợp tổ hợp |
V x D x J
1000 X 15 X 4 |
V x J
300 x 4 |
|
Tổng cộng |
6 x 104 |
1.2 x 103 |
|
|
|
|
Kết hợp tổ hợp |
7.2 x 107 |
|
| |
Những tính toán này không tính đến sự góp
mặt của các chuỗi nhẹ lambda, sự đa dạng đột biến kết hợp soma, chèn khu
vực N hoặc đa đặc hiệu.
Quá trình tái sắp xếp gen của các chuỗi nặng và nhẹ và sự kết hợp tổ hợp
của các chuỗi này xảy ra trong quá trình phát triển tế bào B và không
phụ thuộc vào kháng nguyên. Dòng tế bào B biểu lộ tất cả các tính đặc
hiệu kháng thể có thể, được sản xuất trong quá trình phát triển và kháng
nguyên chỉ cần chọn những dòng mà có các thụ thể thích hợp. Các dòng
được chọn lọc sau đó được hoạt hóa, tăng sinh và biệt hóa thành các tế
bào tương bào sản xuất kháng thể.
THỤ THỂ KHÁNG NGUYÊN CỦA TẾ BÀO
T
Các tế bào T cũng có một thụ thể kháng
nguyên trên bề mặt của chúng. Thụ thể này không phải là một phân tử
kháng thể nhưng nó bao gồm hai chuỗi polypeptid khác nhau mà có các vùng
hằng định và thay đổi tương tự như các kháng thể. Sự đa dạng của các thụ
thể tế bào T cũng được tạo ra cùng một cách như mô tả đối với sự đa dạng
kháng thể (ví dụ bằng cách liên kết VJ và VDJ của các phân đoạn gen và
liên kết tổ hợp). Tuy nhiên, không có đột biến soma đã được quan sát
thấy trong các tế bào T.
|
|
|
 Trở về phần Miễn dịch của Vi khuẩn học và Miễn dịch học
online
Trở về phần Miễn dịch của Vi khuẩn học và Miễn dịch học
online
Page maintained by
Richard Hunt
|
 Hình 2
Hình 2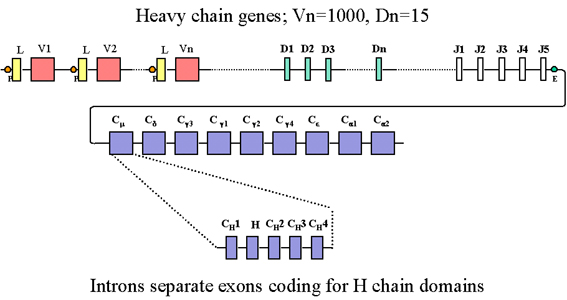 Hình 3 Ngoài một số exon J, gia đình gen chuỗi nặng
cũng có thêm một số exon được gọi là các exon D (đa dạng). Tất cả các
exon được ngăn cách bởi intron
Hình 3 Ngoài một số exon J, gia đình gen chuỗi nặng
cũng có thêm một số exon được gọi là các exon D (đa dạng). Tất cả các
exon được ngăn cách bởi intron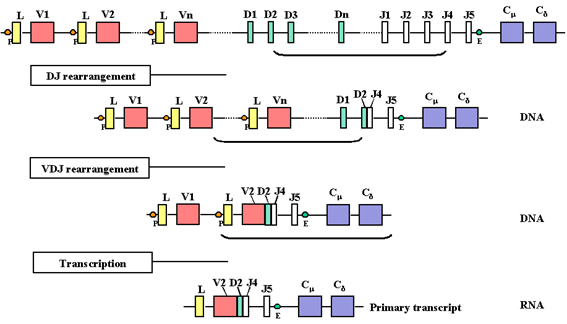 Hình 4
Hình 4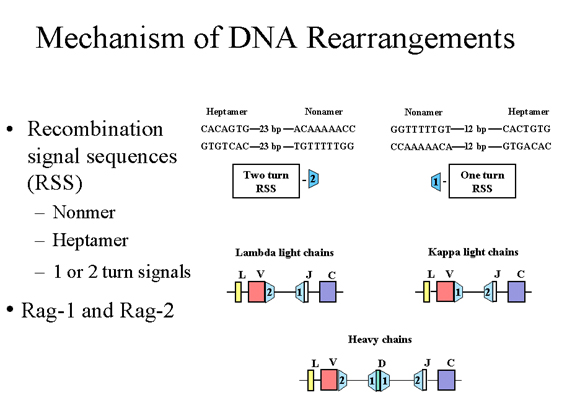 Hình 6
Hình 6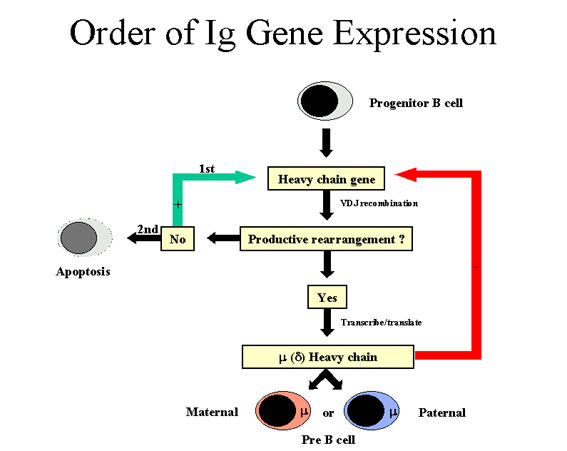 Hình 7
Hình 7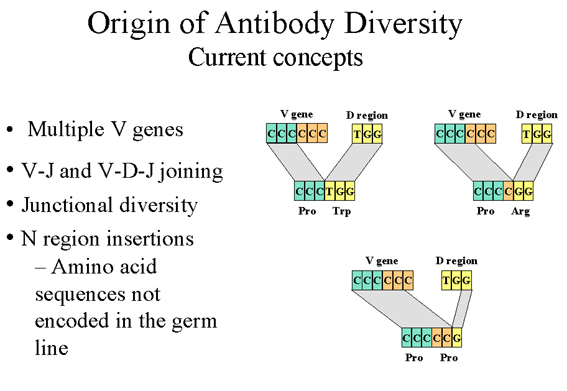 Hình 9 Nguồn gốc các khái niệm hiện nay về sự đa
dạng của kháng thể
Hình 9 Nguồn gốc các khái niệm hiện nay về sự đa
dạng của kháng thể