|
x |
x |
|
 |
 |
|
INFECTIOUS
DISEASE |
BAKTERIYOLOJİ |
IMMUNOLOGY |
MYCOLOGY |
PARASITOLOGY |
VİROLOJİ |
|
ENGLISH |
BAKTERİYOLOJİ - BÖLÜM YEDİ
VİROLOJİ
- BÖLÜM YİRMİDÖRT
BAKTERİYOFAJ
Gene Mayer, PhD
Professor Emeritus
Department of Pathology, Microbiology and Immunology
University of South Carolina School of Medicine
Columbia
Çeviren
Prof. Dr. Mustafa Demirci
İzmir Katip Çelebi Üniversitesi
Tıp Fakültesi, Tıbbi Mikrobiyoloji A.D.
|
|
SPANISH |
|
FRENCH |
|
ALBANIAN |
|
PORTUGUESE |
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|
|

BAKTERIYOLOJİ
Virology

BAKTERIYOLOJİ
Virology
|
|
Logo image © Jeffrey
Nelson, Rush University, Chicago, Illinois and
The MicrobeLibrary |
|
ÖĞRENİM HEDEFLERİ
Bakteriyofajın genel bileşimi ve yapısını açıklamak,
İnfeksiyon süreci ve litik çoğalma döngüsünü tartışmak,
Lizojenik döngü ve düzenlenmesini açıklamak
 © CellsAlive
- James A. Sullivan
© CellsAlive
- James A. Sullivan |
Kompozisyon
Farklı bakteriyofajlar farklı maddeler içeriyor olsalar da hepsi nükleik
asit ve protein içerir. Faja bağlı olarak, nükleik asitleri çeşitli
formlarda olabilir ve her ikisi birlikte olmamak üzere DNA veya RNA
içerirler. Faj nükleik asitleri sık sık olağan dışı veya modifiye
edilmiş bazlar içerirler. Bu modifiye bazlar faj enfeksiyonu sırasında
ev sahibi nükleik asitleri parçalayan nükleazlardan faj nükleik asidini
korur. Nükleik asidin boyutu faja bağlı olarak değişir. Basit fajlar
sadece 3-5 ortalama boyutlu gen ürünleri kodlamak için nükleik aside
sahipken, daha karmaşık fajlar 100'den fazla gen ürünleri için
kodlayabilir.
Faj partikülünde, farklı tür proteinin sayısı ve proteinin her tür
miktarı faja bağlı olarak değişecektir. Basit faj sadece bir ya da iki
farklı tür proteinin birçok kopyası varken, daha karmaşık fajlarda
birçok farklı tür olabilir. Proteinler enfeksiyonda ve çevrede bulunan
nükleazlardan nükleik asitleri koruma işlevi görürler.
Yapı
Bakteriofajlar çok farklı boyutlarda ve şekillerde bulunabilirler.
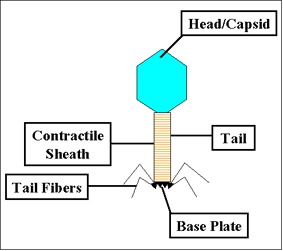
Bakteriyofajların temel yapısal özellikleri, T4 fajının tasvir edildiği, Şekil 1
'de gösterilmiştir.
- Boyut - T4 en büyük fajlar arasındadır; yaklaşık 200 nm uzunluk ve
80-100 nm genişliğindedir. Diğer fajlar daha küçüktür. En çok bulunan
fajların boyutu 24-200 nm uzunluğundadır.
|
|
ANAHTAR KELİMELER
Bakteriyofaj, Faj tipleme, Kapsid, Kuyruk, Kontraktil kılıf, Taban
plakası, Kuyruk lifleri, Öldürücü faj, Tutulma, Erken ve geç m-RNA,
Plaque, Pfu, Lizojeni, Ilıman faj, Profaj, Lizojen, Yapışkan uçlar,
Yer-spesifik rekombinasyon, Baskılanma, İndüksiyon, Lizojenik dönüşüm.

T4 Bacteriophage (TEM x390,000) ©
Dennis Kunkel Microscopy, Inc.
Used with permission
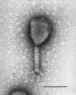
T4 bacteriophage Negative stain electron micrograph
©
ICTV.
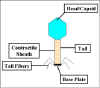 Şekil 1; T4 bakteriyofajın yapısı
Şekil 1; T4 bakteriyofajın yapısı
|
- Kafa veya kapsid - Bütün fajlar, boyutu ve şekli değişebilir bir
baş (kapsid) yapısı içerir. Bazıları ikosahedral (20 taraflı)
diğerleri ipliksidirler. Baş ya da kapsid bir ya da daha farklı
proteinin çok sayıda kopyasından oluşmaktadır. Baş yapısının içinde
nükleik asit bulunur. Baş nükleik asit için koruyucu kaplama olarak
görev yapar.
- Kuyruk - Tüm fajlarda olmamakla birlikte birçok fajda baş
yapısına bağlı kuyruk yapısı vardır. Kuyruk enfeksiyon sırasında
nükleik asidin içinden geçtiği içi boş bir borudur. Kuyruk boyutu
değişebilir ve hatta bazı fajlar bir kuyruk yapısına sahip değildir.
T4 gibi daha karmaşık fajlarda, kuyruk bakterinin enfeksiyonu
sırasında kasılarak nükleik asidi bakteriye aktarma işlevi gören bir
kılıf ile çevrilidir. Kuyruğun sonunda T4 gibi daha karmaşık
fajlarda, bir taban plakası vardır ve buna tutunmuş bir ya da daha
fazla kuyruk lifleri vardır. Taban plakası ve arka lifleri fajın
bakteri hücresine bağlanmasına katılmaktadırlar. Tüm fajların taban
plakaları ve kuyruk lifleri bulunmaz. Bu gibi durumlarda diğer
yapılar faj partikülünün bakteriye bağlanmasında görev almaktadır.
KONAK HÜCRELERİN ENFEKSİYONU
Tutunma (Adsorpsiyon)
Enfeksiyon sürecindeki ilk adım, bakteri hücresi yüzeyine tutunma
basamağıdır. Bu adım kuyruk lifleri veya kuyruk lifleri olmayan
fajlarla bazı benzer yapıların aracılığı ile olmaktadır ve geri
dönüşümlüdür. Kuyruk lifleri bakteride spesifik reseptörlere
bağlanırlar ve fajın konak seçiciliği,(örneğin; bakteriyi enfekte
edebilir.) genellikle kuyruk lifi bulunan fajlarda, kuyruk lifleri
göre belirlenir. Bakteriyel reseptör yapısı farklı bakterilerde
değişir. Bu resöptör yapılarının incelendiği örneklerde bakteride
dış yüzey üzerinde LPS, pili ve lipoprotein içerdiği görülmüştür. Bu
reseptörlerin bakteride başka işlevleri bulunmaktadır ve faj bu
reseptörleri kullanarak enfeksiyon yapmak için gelişmiştir.
Geri dönüşümsüz bağlanma
Kuyruk lifleri aracılığıyla fajın bakteriye bağlanması zayıftır ve
geri dönüşümlüdür. Bir fajın bakteriye geri dönüşümsüz bağlanması
için taban plakası bileşenlerinin bir ya da daha fazlasın olaya
katılması gerekir. Baz plakaları eksik olan fajlarda bakteri
hücresine sıkı bağlanma için başka mekanizmalar vardır.
Kılıf kasılması
Fajın bakteriye geri dönüşsüz bağlanması, kılıfın kasılmasıyla
sonuçlanır (bir kılıfa sahip olan fajlar için) ve içi boş kuyruk
yapısı bakteriyel zarfa (Şekil 2) itilir. Kasılma kılıfları olmayan
fajlar bakteriyel zarfın içine faj parçacığı geçirebilmek için başka
mekanizmalar kullanır. Bazı fajların bakteriyel zarfın çeşitli
bileşenleri sindirmek için enzimleri vardır.
Nükleik Asit Enjeksiyonu
Faj, bakteriyel zarf yoluyla baş kısmında bulunan nükleik asidi içi
boş kuyruk aracılığıyla bakteriye aktarır ve bakteri hücresi içine
girer. Genel olarak, hücre içine giren faj bileşeni sadece nükleik
asididir. Fajın geri kalan yapıları bakterinin dışında kalır. Bu
kuralın bazı istisnaları vardır. Bu virüs partikülünün genellikle
hücre içine alındığı, hayvan hücre virüslerinde farklıdır. Bu fark
muhtemelen, karmaşık yapıları yutmak için, bakterilerin yetersizliği
nedeniyledir.
|
|
MOVIE
Bacteriophage
Requires Quicktime
© Mondo Media
San Francisco, California 94107 USA
and The MicrobeLibrary |
 Şekil 2; T4 kuyruk kılıf kasılması
Şekil 2; T4 kuyruk kılıf kasılması |
FAJ ÇOĞALMA DÖNGÜSÜ
Litik (parçalayıcı) veya Virülan (öldürücü)
Fajlar
Tanım
Litik veya öldürücü fajlar, yalnızca bakteri içinde çoğalma ve fajın
tekrar bakteriyi terk edeceği zaman (yaşam döngüsünün sonunda) bakterini
parçalanması ile hücreyi öldürürler.
Yaşam döngüsü
Litik bir fajın yaşam döngüsü, Şekil 3'te gösterilmiştir.
|
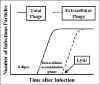 Şekil 3; Litikfajın yaşam döngüsü
Şekil 3; Litikfajın yaşam döngüsü

 Şekil 4; Litik faj deneyi
Şekil 4; Litik faj deneyi |
Tutulma (Eclipse) dönemi
Tutulma aşamasında, hiç bir enfekte faj parçası, bakteri hücresi
dışında yada içinde bulunmaz. Faj nükleik asidi konak biyosentetik
organlarını devralır ve fajın spesifik m-RNA ve proteinleri yapılır.
Fajın, hayvan virüs enfeksiyonlarında göründüğü gibi,
makromolekülerin sentezine yönelik düzenli bir ekspresyonu vardır.
Erken faj m-RNA sı, faj DNA sentezi ve ev sahibi DNA, RNA ve protein
biyosentezinin kapatılması için, gerekli olan erken proteinleri
kodlar. Bazı durumlarda, erken proteinler gerçekte konak
kromozomunun bozulmasına yol açar. Faj DNA yapıldıktan sonra faj
m-RNA yapılır ve faj geç dönem proteinleri yapılır. Geç proteinler
fajın toplanması için gerekli olduğu gibi bakteriyel hücre
parçalanması için gerekli yapısal proteinlerdir.
Hücre içi birikim fazı
Bu aşamada, yapılan nükleik asit ve yapısal proteinler toplanır ve
bulaştırıcı faj partikülleri hücre içinde birikir.
Lizis ve salınma fazı
Bir süre sonra bakteriler, ortama salınan faj litik proteinleri ve
hücre içi faj birikimi nedeniyle, parçalanmaya başlar. Enfekte
edilen bakteri başına salınan faj taneciklerinin sayısı 1000 kadar
yüksek olabilir.
Litik fajın test edilmesi
Agar pleyt deneyi
Litik faj bir agar pleyt deneyi ile belirlenebilir. Üzerine faj ve
bakteri ekilmiş agar pleytinin temiz olması bakterilerin lizise
uğradığını gösterir (Şekil 4). Her temiz plak bir bulaşıcı fajdan
büyür. Her bakterinin üremediği alana (plak) yol açan infeksiyöz faj
parçacığı, bir pfu (plak oluşturma birimi) olarak adlandırılır.
|
 Şekil 5; Faj kromozomun daire haline gelmesi: yapışkan uçlar
Şekil 5; Faj kromozomun daire haline gelmesi: yapışkan uçlar
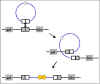 Şekil 6; Bölgeye özgü rekombinasyon
Şekil 6; Bölgeye özgü rekombinasyon |
Lizojenik veya ılımlı faj
Tanım
Lizojenik veya ılıman fajlar hem litik döngü yoluyla çoğalabilirler hem
de hücrede suskun bir duruma girebilirler. Bu suskun (uyur) halde
bulunan faj genlerinin çoğunda transkripsiyon olmaz; faj genomu
bastırılmış bir durumda bulunmaktadır. Bu durumda ortada bir faj
olmadığı fakat bir fal üretme potansiyeli olduğu için, bastırılmış halde
bulunan faj DNA'sına profaj denir. Çoğu durumda faj DNA'sı gerçekten
konak kromozomuna bütünleşmiş olur ve konak kromozomu ile birlikte
çoğalır ve yavru hücrelere aktarılır. Bir profaj barındıran hücre
profajının varlığından olumsuz etkilenmez ve lizojenik durum süresiz
devam edebilir. Bir profaj barındıran hücre lizojen olarak adlandırılır.
Lizojeniye yol açan olaylar
Prototip Faj: Lambda
Faj kromozomun daire halinde olması
Lambda DNA, 5 'ucunda küçük tek iplikçikli bölgeleri de içeren iki
iplikçikli doğrusal bir moleküldür. Bu tek iplikçikli uçlar
tamamlayıcıdırlar (yapışkan uçlar) böylece baz çifti ve dairesel
molekül üretilebilir. Hücrede dairenin serbest uçları, Şekil 5'te
gösterildiği gibi, kovalent olarak kapalı bir daire oluşturacak
şekilde bağlanabilir.
|
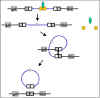 Şekil 7; Lizojeninin sonlandırılması
Şekil 7; Lizojeninin sonlandırılması
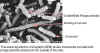 Şekil 8A; Hücrelerin dış kısmında (küçük beyaz noktalar gibi) faj
partikülleri ile birlikte Escherichia coli hücrelerinin tarama elektron
mikrografıdır (SEM). © ScottKachlany, Cornell
UniversityIthaca, New York, ABD ve MicrobeLibrary
Şekil 8A; Hücrelerin dış kısmında (küçük beyaz noktalar gibi) faj
partikülleri ile birlikte Escherichia coli hücrelerinin tarama elektron
mikrografıdır (SEM). © ScottKachlany, Cornell
UniversityIthaca, New York, ABD ve MicrobeLibrary
 Şekil 8B;
Şekil 8B;
SEM, muhtemelen faj salınması nedeniyle parçalanan hücre zarfı ile E.
coli hücreleri. Faj konak hücre içinde çoğaldığı zaman, konak hücreden
serbest bırakılması gerekir. Bu genellikle hücrenin parçalanmasyla
meydana gelir. © ScottKachlany, Cornell UniversityIthaca,
New York, ABD ve MicrobeLibrary
|
Bölgeye özgü rekombinasyon
Bir fajın kodladığı enzim tarafından katalize edilen bir
rekombinasyon olayı, daireselleşmiş faj DNA üzerinde belirli bir yer
ile ana kromozom üzerinde belirli bir yer arasında gerçekleşir.
Şekil 6'da gösterildiği gibi sonuç konak kromozomuna faj DNA
entegrasyonudur.
Faj genomunun bastırılması (represyonu)
Represör olarak adlandırılan, bir faj tarafından kodlanan protein,
faj DNA üzerinde, operatör denilen, belirli bir yere bağlanır ve
represör geni HARİÇ bir çok faj geninin transkripsiyonunu kapatır.
Sonuçta, konakçı kromozomuna entegre edilmiş, bir sabit bastırılmış
faj genomu olur. Her ılıman faj sadece kendi DNA'sını
baskılayabildiği ve diğer fajları baskılayamadığı için represyon çok
özeldir (aynı faj ile süper enfeksiyona bağışıklık).
Lizojeninin sonlandırılmasına yol açan olaylar
Herhangi bir zamanda bir lizojenik bakteri olumsuz koşullara maruz
kaldığında, lizojenik durum sona erdirilebilir. Bu süreç indüksiyon
olarak adlandırılır. Lizojeninin sona erdirilmesine yol açan durumlar;
kuruma, UV maruziyeti veya iyonize radyasyon, mutajenik kimyasallara
maruz kalma v.b. gibi olumsuz koşullarda, represör proteini imha eden,
proteazların (rec A protein) üretimine yol açar. Bu da, entegrasyon
sürecinin tersine çevrilmesi ve litik çoğalma faj genlerinin
ekspresyonuna yol açmaktadır. Şekil 7.
Litik veya Lizojenik Döngü
Lambda fajının litik veya lizojenik döngüye girme kararı, bir hücreye
ilk girdiğinde hücre represör ve hücrede cro adı verilen başka bir faj
proteini konsantrasyonu ile belirlenir. Cro proteini represör sentezini
kapatır ve böylece lizojeninin oluşmasını engeller. Cro üretimini
destekleyen çevresel koşullar litik döngüye yol açarken, represör
üretimini destekleyenler lizojenik döngünün oluşmasına yol açacaktır.
Lizojeninin önemi
Hayvan virüs transformasyonu için model
Lizojeni hayvan hücrelerinin virüs transformasyonu için bir model
sistemdir
Lizojenik dönüşüm
Hücre lizojenik olduğu zaman, faj hücrede exprese olduğunda bazen
ilave genler taşınır. Bu genler bakteri hücresinin özelliklerini
değiştirebilir. Bu süreçe lizojen veya faj dönüşümü denir. Bu klinik
öneme sahip olabilir. Örneğin, Lizojenikfajların, bağışıklığı
etkileyen önemli antijenlerden biri olan, Salmonella O antijeni
yapısını değiştirebilecek genleri taşıyabileceği gösterilmiştir.
Corynebacterium diphtheriae'da toksin üretimine, bir faj tarafından
taşınan bir gen aracılık eder. Yalnızca faj ile lizojen olan suş
patojenikdir.
|
|
|
 Mikrobiyoloji ve İmmünoloji On-line Kitabı Bakteriyoloji Bölümüne
dön
Mikrobiyoloji ve İmmünoloji On-line Kitabı Bakteriyoloji Bölümüne
dön
Mikrobiyoloji ve İmmünoloji On-line, Viroloji Bölümüne Dönünüz
This page last changed on
Wednesday, November 23, 2016
Page maintained by
Richard Hunt
|
 Şekil 2; T4 kuyruk kılıf kasılması
Şekil 2; T4 kuyruk kılıf kasılması Şekil 3; Litikfajın yaşam döngüsü
Şekil 3; Litikfajın yaşam döngüsü Şekil 7; Lizojeninin sonlandırılması
Şekil 7; Lizojeninin sonlandırılması