| x | x | ||||
|
|
 |
||||
| BAKTERIYOLOJİ | İMMÜNOLOJİ | MYCOLOGY | PARASITOLOGY | VIROLOGY | |
|
|||||
| ENGLISH | |||||
| EN FRANCAIS | |||||
| EM PORTUGUES - PORTUGUESE | |||||
|
Let us know what you think FEEDBACK |
|||||
| SEARCH | |||||
|
|
|||||
| Logo image © Jeffrey Nelson, Rush University, Chicago, Illinois and The MicrobeLibrary | |||||
|
ÖĞRENİM HEDEFLERİ
ANAHTAR KELİMELER |
B HÜCRE RESPTÖRÜ ve T
HÜCRE RESEPTÖRÜ KARŞILAŞTIRILMASI B hücreleri ve T hücreleri, antijen olarak farklı maddeleri farklı biçimde tanır. B hücresi reseptör olarak, hücre yüzeyine bağlı immünoglobülini kullanır ve bu aktivasyonundan sonra salgılanan imünoglobulin ile bu reseptörün özgünlüğü aynıdır. B hücreleri çözünebilir formdaki aşağıdaki antijenleri tanır:
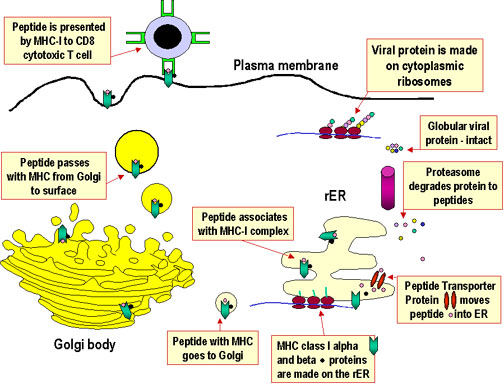
Antijen işlenmesi ve sunumu hücrede, proteinlerin parçalanması (proteoliz), MHC molekülleri ile bu parçaların etkileşimi ve hücre yüzeyinde T hücre yüzeyindeki T hücre reseptörü tarafından tanınabilen peptit-MHC molekülünün ekspresyonuyla sonuçlanan proseslerdir. Ancak, protein parçalarının MHC molekülleriyle etkileşimine sebep olan yolaklar sınıf I ve II MHC için farklıdır. MHC sınıf I molekülleri sitosoldeki hücre içi (endojen) proteinlerin parçalanma ürünlerini sunar. MHC sınıf II molekülleri hücre içi kompartmanlarında yer alan ekstraselüler (ekzojen) proteinlerden oluşmuş fragmentleri sunar.
|
||||
|
|
|
||||
 Şekil 2
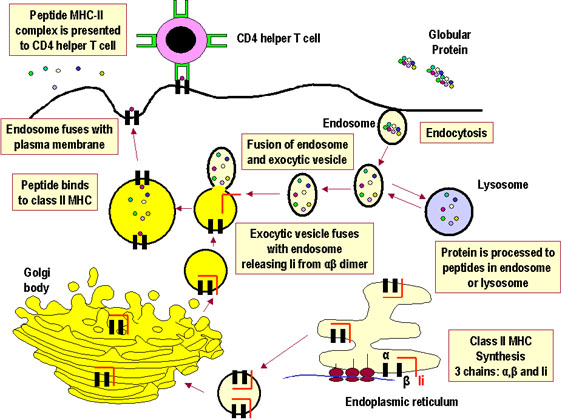
Şekil 2MHC-II ile kısıtlı ekzojen antijen sunum yolağı |
|||||
|
|
|
||||
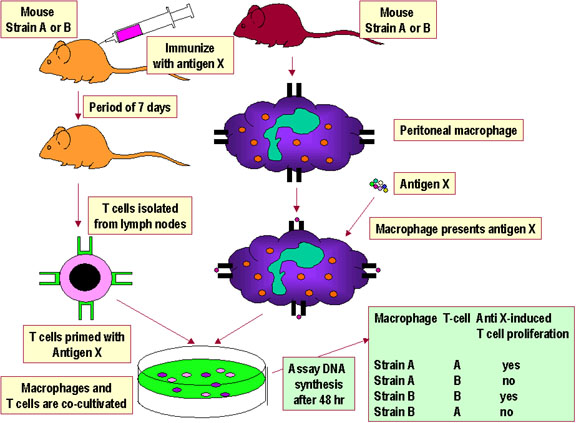 Şekil 3 Th/APC etkileşimlerinin
self MHC kısıtlaması
Şekil 3 Th/APC etkileşimlerinin
self MHC kısıtlaması |
|||||
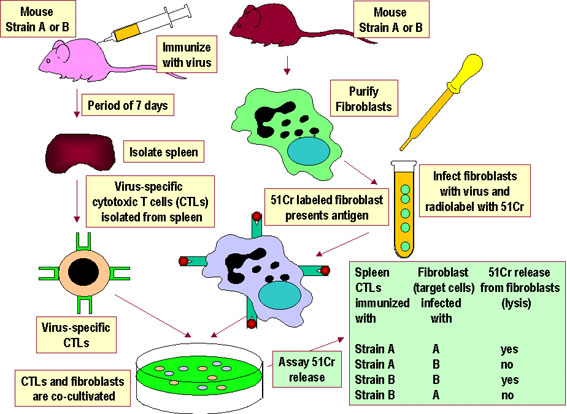 Şekil 4
Şekil 4A veya B suşu fareden virüs-spesifik CTL’ler sadece spesifik virüsle enfekte sinjenik hedef hücrelerini lize ederler. CTL’ler enfekte olmayan hedef hücreleri lize etmezler ve alloreaktif değildirler. Daha sonraki analizler, CTL’lerin ve hedef hücrelerin viral antijenleri CTL’lere sunmak için sınıf I MHC allellerini paylaşan hayvanlardan gelmek zorunda olduğunu göstermiştir. |
SELF MHC KISITLAMASI T hücresinin yabancı bir protein antijeni tanıması ve buna karşı yanıt oluşturması için antijeni sunan hücrenin yüzeyindeki MHC’yi öz MHC olarak tanıması gerekir. Bu, self MHC kısıtlaması olarak adlandırılmaktadır. Yardımcı T hücreleri kendi sınıf II MHC varlığında antijeni tanır. Sitolitik T hücreleri kendi sınıf I MHC varlığında antijeni tanır. T hücrelerinin kendi MHC moleküllerini tanıması için sınırlanmış hale gelmesiyle gerçekleşen bu proses timusta gerçekleşir. ASH-yardımcı T hücre etkileşimi ve sınıf I MHC-sitotoksik T hücre
etkileşimi için self MHC kısıtlaması gösteren deneysel sistemler sırasıyla
Şekil 3 ve 4’te gösterilmiştir. |
||||
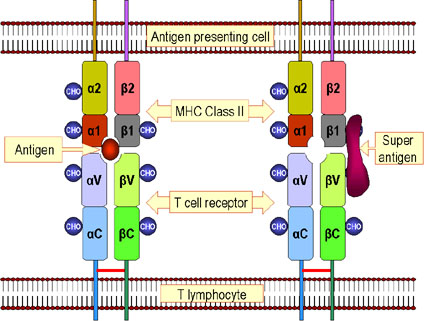 Şekil 5
Şekil 5Antijen ve süperantijen arasındaki farklılılar. Antijenik peptitler hücrede işlenir ve hücre yüzeyinde sınıf II MHC molekülleriyle etkileşerek sunulur. Daha sonra bunlar T hücre yüzeyindeki T-hücre reseptörünü tetikler. Süperantijenler işlenmezler ancak sınıf II MHC proteinine ve T hücre reseptörünün V beta zincirine bağlanırlar. Verilen süperantijen, belirli bir V beta zinciri eksprese eden farklı bir T hücre sınıfını aktive eder. Not: MHC-II-TCR’nin normal bir şekilde işlenmiş peptit etkileşimi durumunda MHC molekülü üzerindeki peptidin tanınması TCR’nin V alfa, J alfa, V beta, D beta ve J beta segmentlerine ihtiyaç duyar.Bu tip bir etkileşim çok az oranda oluşur. MHC II-TCR’nin işlenmemiş bir süperantijenle etkileşimi durumunda sadece belirli bir V beta bölgesi tanınır. Bu daha fazla sıklıkta oluşmaktadır. |
ANTİJEN SUNAN HÜCRELER
Antijen sunan hücrelerin üç temel tipi dendritik hücreler, makrofajlar ve B
hücreleri olup sınıf II MHC moleküllerini eksprese eden diğer hücreler de (ör:
timik epitelyal hücreler) bazı durumlarda antijen sunan hücreler olarak rol
oynar. Deri ve diğer dokularda bulunan dendritik hücreler antijenleri
pinositozla sindirir ve antijenleri lenf nodlarına ve dalağa taşır. Lenf
nodlarında ve dalakta çoğunlukla T hücre bölgelerinde bulunurlar. Dendritik
hücreler en etkili antijen sunan hücrelerdir ve antijenleri naif (virjin) T
hücrelerine sunarlar. Bunun ötesinde içselleşmiş antijenlerin yolağı
çoğunlukla sınıf II yolağı olmasına rağmen bunlar içselleşmiş antijenleri
hem sınıf I hem de sınıf II MHC molekülleriyle etkileşime girerek
sunabilirler. İkinci antijen sunan hücre tipi ise makrofajlardır. Bu
hücreler antijenleri fagositoz veya pinositozla sindirirler. Makrofajlar
naif T hücrelerine antijen sunumunda çok etkili değildirler ancak hafıza T
hücrelerini aktive etmede çok iyidirler. Antijen sunan hücrelerin üçüncü
tipi B hücreleridir. Bu hücreler antijene yüzey immünoglobulinleri ile
bağlanırlar ve pinositozla sindirirler. Makrofajlar gibi bu hücreler de
antijenlerin naif T hücrelerine sunumunda dendritik hücreler kadar etkili
değildirler. B hücreleri antijenin hafıza T hücrelerine sunumunda çok
etkilidirler, özellikle antijen konsantrasyonu çok düşük olduğunda
etkililerdir çünkü B hücrelerinin yüzeyindeki immünoglobulinler antijene
yüksek afiniteyle bağlanır.
Süperantijenler, patolojik etkiye sahip olabilen çok miktarda sitokin
üretmesi yol açan T hücrelerini poliklonal olarak aktive edebilen
antijenlerdir. Bu antijenler T hücrelerine sınıf II MHC molekülleriyle
etkileşerek sunulmalıdır ancak antijenin işlenmesine gerek yoktur. Şekil 5
konvansiyonel antijenlerin ve süperantijenlerin T hücrelerine nasıl
sunulduğunu karşılaştırmaktadır. Bir süperantijen varlığında işlenmemiş
protein sınıf II MHC molekülüne ve TCR’nin bir veya daha fazla Vβ
bölgesine bağlanır. Bu antijen MHC molekülünün peptit bağlama çukuruna veya
TCR’nin antijen bağlama bölgesine bağlanmaz. Bu yüzden, TCR’de belirli bir Vβ
kullanan herhangi bir T hücresi süperantijen tarafından aktive olacaktır ve
çok sayıda T hücrein aktivasyonuyla sonuçlanacaktır. Her bir süperantijen
farklı Vβ bölgesine bağlanacaktır. |
||||
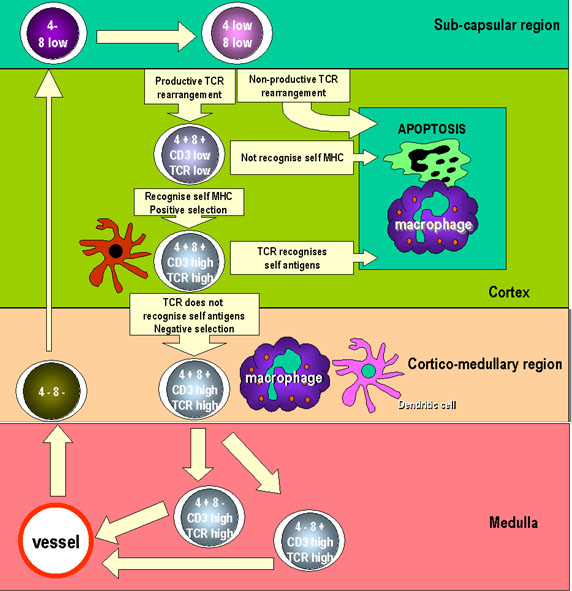 Şekil 6
Şekil 6Pretimik T hücreleri timus rudimentine girer ve timusun subkapsüler bölgesinde büyük lenfoblastlar olarak çoğalırlar. Bu lenfoblastlar çoğalarak farklılaşan hücre havuzlarına neden olurlar. Burada hücreler CD8 ve CD4 pozitif hale gelirler ancak ekspresyonları azdır. TCR genleri aynı zamanda bu hücrelerde yeniden düzenlenir ve ürünler de hücre yüzeyinde az miktarda eksprese edilebilir. Hücreler olgunlaşınca uzun ve dallanmış olan kortikal epitelyal hücrelere bağlandıkları yer olan kortekse hareket ederler ve diğer hücrelerle etkileşime girmek için geniş bir alan oluştururlar. Timositlerin yüzeyindeki TCR’ler epitelyal hücrelerin yüzeyindeki MHC molekülleriyle etkileşime girerek pozitif seleksiyona sebep olur. Seçilmeyen hücreler apoptoza gider ve makrofajlar tarafından fagosite edilirler. Timositler timusun korteksine göç ettiklerinde CD3, CD4, CD8 ve TCR ekspresyonu artar. Dendritik hücrelerin ve makrofajların sunduğu otoantijenlerle etkileşime girdiğinden kendine karşı reaktivite gösteren hücreler yok edilir. Bu, negatif seleksiyona sebep olur. Cells CD4 veya CD8 eksprese eden hücreler oluşur ve kortiko-medüllar bölgede özelleşmiş damarlarla perifere göç ederler. |
TİMİK EĞİTİM Th ve Tc hücrelerinin ikisi de
self-MHC sınırlıdır. Ayrıca, T hücreleri normalde kişinin kendi
antijenlerini tanımaz. Peki self MHC sınırlı T hücreleri nasıl oluşur ve
kendimize reaktif olan T hücreleri neden üretilmez? T hücrelerindeki
rastgele VDJ düzenlemelerinin bazı kendinden olmayan MHC’leri ve bazı kendi
antijenlerini tanıyan T hücrelerini oluşturması beklenmektedir. Sadece
perifere giden T hücrelerin self-MHC sınırlı olduklarından ve kendi
antijeniyle etkileşime giremediklerinden emin olmak timusun görevidir.
Periferdeki fonksiyonel T hücreleri self MHC ile etkileşime giren yabancı
antijenleri tanımalıdır çünkü ASH veya hedef hücreler kendi MHC’si ile
etkileşimde olan yabancı antijeni sunar. Ancak, kişinin yabancı MHC ile
etkileşimde olan antijeni (kendinin veya yabancı) tanıyan fonksiyonel T
hücrelerine periferde ihtiyacı yoktur. Kişi, özellikle kendi MHC’si ile
etkileşimde olan kendi antijenini tanıyan fonksiyonel T hücrelerini
periferinde istemez çünkü bunlar sağlıklı, normal dokularda hasara sebep
olabilir. Timustaki pozitif ve negatif seçilim %100 etkili bir süreç değildir. Ayrıca, timusta bütün kendi antijenleri eksprese edilmiyor olabilir. Bu yüzden, bazı kendine reaktif T hücreleri perifere gidebilir. Bundan dolayı da periferdeki kendine reaktif T hücrelerini elimine etmek için tasarlanmış ek mekanizmalar bulunmaktadır. Bu kısım tolerans bölümünde tartışılacaktır. B HÜCRE SEÇİMİ B hücreleri MHC kısıtlı
olmadığından B hücreleri için pozitif seleksiyona gerek yoktur. Ancak B
hücrelerin de negatif seleksiyonu (ör: self reaktif klonların ortadan
kaldırılması) gereklidir. Bu, kemik iliğinde B hücre gelişimi esnasında
gerçekleşir. Ancak, B hücrelerinin negatif seçilimi T hücrelerininki kadar
kritik değildir, çoğu örnekte, B hücreleri aktif hale gelmek için T
hücrelerinin yardımına ihtiyaç duyar. Bu yüzden, eğer bir self reaktif B
hücresi perifere geçerse T hücre yardımı olmadığından aktive olamayacaktır. |
||||
|
|||||
|
|
|||||
|
|
|||||
|
|
|
||||