|
x |
x |
|
 |
 |
|
INFECTIOUS
DISEASE |
BAKTERIYOLOJİ |
İMMÜNOLOJİ |
MYCOLOGY |
PARASITOLOGY |
VİROLOJİ |
|
ENGLISH |
VIROLOJİ - BÖLÜM YEDİ
KISIM ON
İNSAN İMMUNYETMEZLİK VIRUSU VE AIDS
CD4 HÜCRELERİN KAYBI
Dr Richard Hunt
Professor
Department of Pathology, Microbiology and Immunology
University of South Carolina School of Medicine
Çeviren. Prof. Dr. Selçuk
Kaya
İzmir Katip Çelebi Üniversitesi, Tıp Fakültesi
|
|
En
Español |
|
SHQIP - ALBANIAN |
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|

 |
|
BU BÖLÜM ÇEŞİTLİ KISIMLARINDA SONRAKİ BÖLÜME GİTMEK İÇİN YUKARIDAKİ NEXT >>
düğmesini kullanın
VİROLOJİ SEKİZİNCİ BÖLÜME DEVAM İÇİN TIKLAYINIZ
BURAYA
|
|
DİĞER HIV VE AIDS BÖLÜMLERİ İÇİN LİNKLER BU sayfanın
alt kısmında |

Tomurcuklanma hücre lizisine neden olur
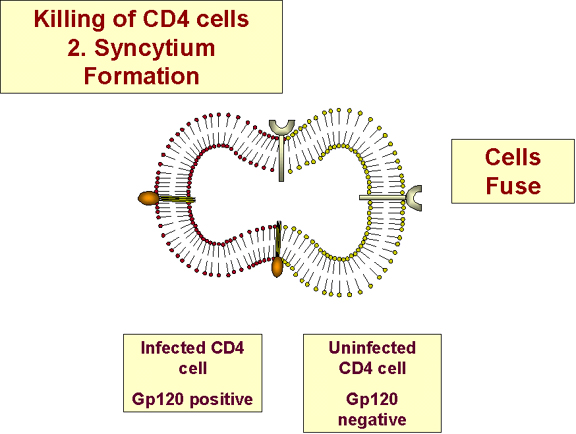
 Sinsitya oluşumu
Sinsitya oluşumu
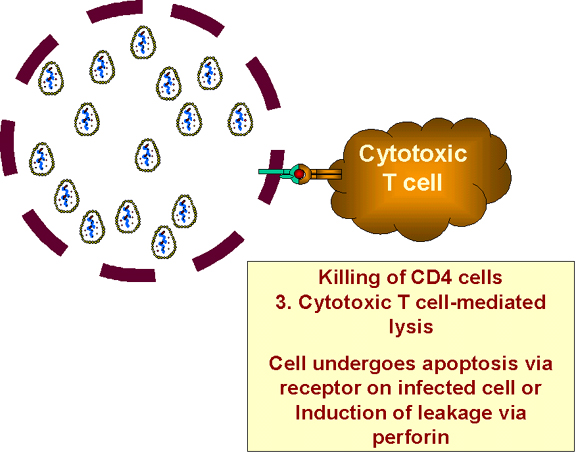
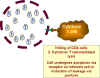 Enfekte
hücreler, sitotoksik T-hücreleri tarafından yok edilir Enfekte
hücreler, sitotoksik T-hücreleri tarafından yok edilir
Şekil 27 - HIV enfeksiyonu sonrası T4 hücrelerinin kaybı için bazı
olası mekanizmalar
|
NEDEN CD4 + YARDIMCI T
HÜCRELERİNİN İLERLEYEN BİR KAYBI VAR?
NEDEN CD8 + KATİL T HÜCRELERİ HASTALIĞIN İLERİ EVRELERİNDE YOK OLUR?
Sadece 1/ 10.000 (erken) veya 40 de 1 (daha sonra) hücreler
üretken enfeksiyon gösterirken neden T4 hücrelerinin tümü kaybolur? CD4 +
hücrelerinin neden tümü kaybolur hala belirsiz ancak olasılıklar vardır:
-
Enfekte CD4 hücre aktive olduğunda, çok sayıda virion
sentezlenir. Bunlar hücreden tomurcuklanır ve delinmiş membran ile
sonuçlanır (Şekil 27). Ama bunun için hücrenin enfekte olması gerekir ve
çoğu CD4 hücreleri enfekte değildir.
-
HIV'in zarı ile hücrenin membranı pH 'dan bağımsız bir
mekanizma ile enfekte edilmesi ile birleşebildiğinden, sinsitia formasyonu
enfekte olmamış hücrelere virüsün yayılmasına neden olabilir (Şekil 27). Ama
sinsitia çok yaygın değildir.
-
Viral proteinleri üreten enfekte hücreler (latent olanlar
değil), sınıf I MHC histokompatibilite antijenlerine ile bağlantılı olarak
hücre yüzeyi üzerinde bu proteinleri sunacaktır. Enfekte hücre, diğer viral
enfeksiyonlu hücreler gibi, sitotoksik T hücreleri tarafından yok edilir (Şekil
27). Yine bu durum, HIV ile enfekte olmuş hücrelerde olur.
-
gp120, kovalent olmayan etkileşimler ile virüs yüzeyinde
gp41 ile bağlantılıdır ve sıklıkla enfekte hücrelerden ya da virüs
parçacıklarından saçılır. Bu CD4 antijen ile enfekte olmamış hücrelere
bağlanır. Bunun bir sonucu olarak, enfekte edilir gibi görünmektedir ve
bağışıklık sistemi tarafından yok edilir.
-
Aktif fakat enfekte olmamış T4 hücrelerinin yüzeyinde
spesifik bir antijen ile reaksiyona girebilecek enfeksiyonlu hastalarda AIDS
ile ilişkili sitotoksik antikorlar rapor edilmiştir.
|
| |
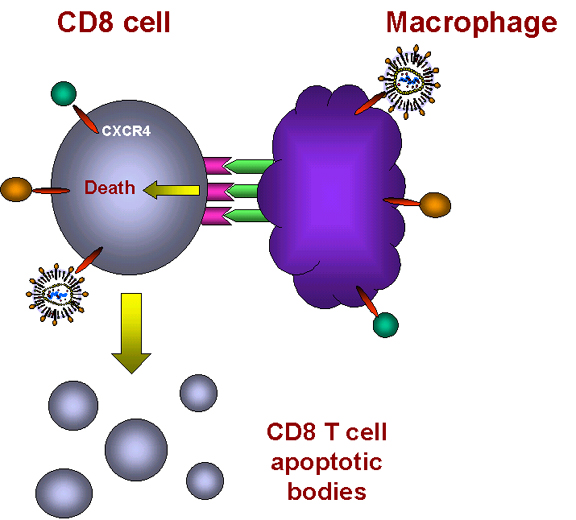
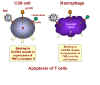 Sitokin
bağlanmasınmakrofajda TNFalfa ekspresyonu ve CD8 + T hücresinde reseptör
ekspresyonunu indükler Sitokin
bağlanmasınmakrofajda TNFalfa ekspresyonu ve CD8 + T hücresinde reseptör
ekspresyonunu indükler
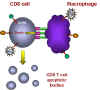 Hücreler başka biri temas eder ve TNF-alfa ve reseptör etkileşim olur. apoptozu
meydana gelmektedir
Hücreler başka biri temas eder ve TNF-alfa ve reseptör etkileşim olur. apoptozu
meydana gelmektedir
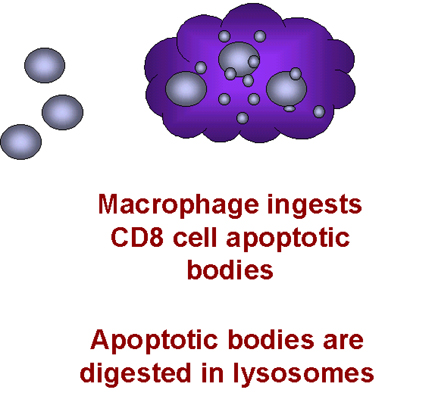
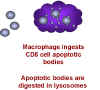 Makrofajlar T hücrelerini içselleştirir
Makrofajlar T hücrelerini içselleştirir
Şekil 28 - T8 hücrelerinde apoptoz indüklenmesi
|
-
AIDS'in bir oto-immün bileşenini olabilir. Normalde
antijenik yanıt T4 hücreleri tarafından yürütülür, CD4 antijeni MHC tip II
doku uygunluk antijenleri ile etkileşime girer. GP120 ayrıca CD4
bağlandığından hem de CD4-bağlama bölgesi olduğundan, GP120 MHC sınıf II
antijenlerini taklit edebilir; Gerçekten de, iki protein içinde benzer
sekans bölgelerinin olduğu görülmektedir. Böylece, anti-gp 120 antikorları
yanı sıra, anti-MHC antikorları ortaya çıkabilir. (Bu aşı üretimi için sorun
teşkil edebilir).
-
HIV T4 hücrelerinin bir alt kümesini enfekte etmesi
mümkündür bu T4 hücrelerinin tüm nüfusunun yayılımı için hayati önem
taşımaktadır
-
HIV proteinleri T4 hücre fonksiyonunu değiştirebilir. Bunun
için bazı kanıtlar vardır.
-
Halen, bütün CD4 + ve CD8 + hücre popülasyonunun kaybı için
en aktif çalışılan olasılık, HIV (Bu apoptoz otoimmün üstesinden gelmek ve
bir bağışıklık tepkisini sonlandırmak için T4 hücrelerinde normal bir
süreçtir), bu hücrelerde apoptozu başlatır olmasıdır (Şekil 28). Bu,
hastalık ilerlemesi sırasında CD4 hücre kaybında önemli bir faktör olduğu
düşünülmektedir
T4 hücrelerin sadece bir azınlığın belirli bir zamanda enfekte
olması ve hastalığın ileri evrelerinde tümünün kaybolur görünmesinin nedenlerini
yukarıda bazıları açıklayabilir. Ayrıca onun ko-receptor geçişi gibi virüs başka
bir T4 hücre popülasyonu geçişi olabilir (yukarı bakınız).
CD8 + hücrelerinin sadece küçük sayıları HIV ile enfekte edilir
ve onların seviyeleri uzun yıllar hastalığın seyri sırasında yüksek kalır. Ve
sonra, son zamanlarında açıklanamaz şekilde hızla ölür. Enfeksiyonda geç oluşan
HIV alt tiplerinin bazılarında CD8 hücrelerinin kitlesel apoptozu istemi ortaya
çıkmaktadır. CD8 hücreleri, çoğunlukla CD4- olmasına rağmen, CXCR4 ko-reseptörü
var ve HIV buna bağlanabilir (HIV'in sadece daha sonra sinsityum oluşturan
suşları). Küçük CD4 antijen mevcut olduğu için hiçbir enfeksiyon olmaz ancak
CXCR4'e bağlanma hücreye bir sinyal gönderir, apoptoz ve kütle CD8 + hücresi
intiharı için bir sinyal meydana gelmektedir.
|
|
|
Bu nasıl oluyor? Şimdi bilinmektedir ki, HIV suşlarının
bağlanması tümör nekroz-alfa ölüm iletim yolu hareketinde CXCR4 reseptör
setlerine daha sonra enfeksiyonu ulaştırır (Şekil 28). Makrofajlarda, hücre
yüzeyi üzerinde CXCR4 reseptörüne bağlanmak için bir ligand olan TNF-alfa
ekspresyonunu teşvik eder. CD8 + T hücrelerinde, aynı bağlanma TNF-a reseptör II
ifadesini tetikler.
Böyle bir makrofaj ve CD8 + T hücre temas ettiğinde, makrofajda
TNF-a, CD8 + T hücresi üzerindeki TNF-a reseptörüne bağlanmaktadır. Bu, CD8 + T
hücresinin vezikülasyonunda elde edilen CD8 + T hücresinde bir apoptoz sinyali
tetikler (Şekil 28). Makrofajlar sonra T hücresinin kalıntılarını fagosite eder.
Bu Makrofajların CD8 + hücrelerinin ölmesi için neden mevcut olması gerektiğinin
açıklıyor. Neden bu doğal olarak olur? Neden kemokinler, CD8 + T hücreleri için
ölüm sinyalleri olarak hareket eder? Bu hücreler katil hücrelerdir ve onlar
yanlış yerde sonuçlanırsa ciddi sorunlara yol açabilir. Onlar uygun konuma
ulaşmadıkça kemokinler, makrofaj-aracılı ölümün kaderine doğrudan CD8 + T
hücreleri olduğu düşünülmektedir.
|
|
|
|
|
|
HIV’IN DİĞER KISIMLARI
KISIM I İNSAN IMMUNYETMEZLİK VIRUS VE AIDS
KISIM II HIV VE AIDS HASTALIĞI
KISIM III HASTALIĞIN GİDİŞATI
KISIM IV İLERLEME VE KOFAKTORLER
KISIM V İSTATISTIK
KISIM VI SUBTİPLER VE KO-RESEPTORLER
KISIM VII HIV’IN HAYAT SİKLUSU VE
KOMPONENTLERİ
KISIM VIII HIV’IN LATENLİĞİ
KISIM IX HIV GENOMU
KISIM X CD4 HÜCRE KAYBI
KISIM XI POPULASYON POLİMORFIZMİ
EK I ANTI-HIV AŞILARI
EK II HIV AIDS’E Mİ NEDEN OLUR?
EK III ANTI-HIV KEMOTERAPİSİ
|
| |
|
| |
 Mikrobiyoloji ve İmmünoloji On-line, Viroloji Bölümüne Dönünüz
Mikrobiyoloji ve İmmünoloji On-line, Viroloji Bölümüne Dönünüz
This page last changed on
Sunday, August 28, 2016
Page maintained by
Richard Hunt
|

 Sitokin
bağlanmasınmakrofajda TNFalfa ekspresyonu ve CD8 + T hücresinde reseptör
ekspresyonunu indükler
Sitokin
bağlanmasınmakrofajda TNFalfa ekspresyonu ve CD8 + T hücresinde reseptör
ekspresyonunu indükler