| x | x | ||||
|
|
 |
||||
| VI KHUẨN HỌC | MIỄN DỊCH HỌC | NẤM HỌC | KÝ SINH TRÙNG HỌC | VIRÚT HỌC | |
| ENGLISH |
|
||||
| TURKISH | |||||
| FRANCAIS | |||||
| PORTUGUES | |||||
|
Let us know what you think FEEDBACK |
|||||
| SEARCH | |||||
|
|
|||||
| Logo image © Jeffrey Nelson, Rush University, Chicago, Illinois and The MicrobeLibrary | |||||
|
MỤC TIÊU GIẢNG DẠY
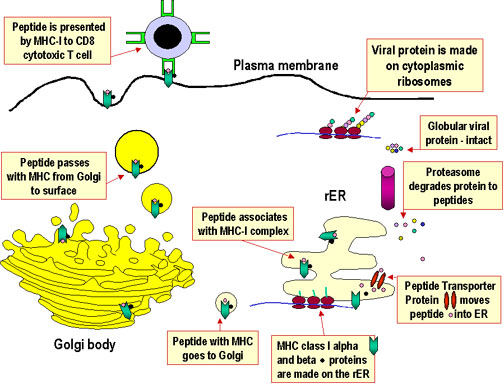
Kháng nguyên nội sinh
|
SO SÁNH BCR VÀ TCR Các tế bào B và T nhận biết nhiều kháng nguyên khác nhau với những cách thức khác nhau. Các tế bào B sử dụng kháng thể bề mặt tế bào như là một thụ thể và đặc hiệu tương tự như các kháng thể mà nó tiết ra sau khi hoạt hóa. Tế bào B nhận biết các kháng nguyên sau đây ở dạng hòa tan:
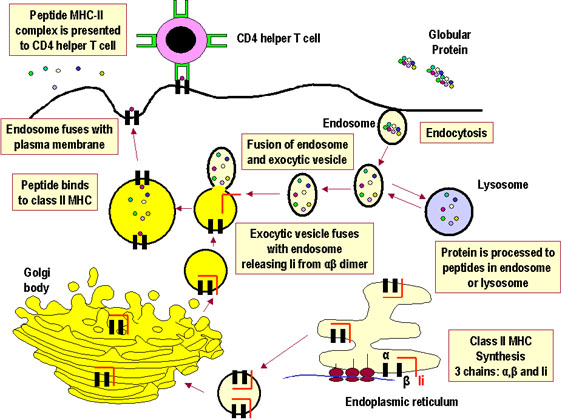
Sự xử lý và trình diện kháng nguyên là một quá trình xảy ra bên trong tế bào dẫn đến phân cắt (sự phân giải protein) các protein, kết hợp các mảnh protein với các phân tử MHC, và biểu lộ của các phân tử peptid-MHC ở bề mặt tế bào, nơi chúng có thể được nhận biết bởi các thụ thể nằm trên một tế bào T. Tuy nhiên, con đường hình thành sự kết hợp của các đoạn protein với các phân tử MHC khác nhau đối với MHC lớp I và II. Các sản phẩm của phân tử MHC lớp I trình diện các sản phẩm thoái hóa có nguồn gốc từ protein trong bào tương của tế bào (nội sinh). Các phân tử MHC lớp II trình diện các mảnh peptid có nguồn gốc từ các phân tử protein ngoại (ngoại sinh) được đặt trong một khoang nội bào.
|
||||
|
|
|
||||
 Hình 2
Hình 2Con đường trình diện kháng nguyên ngoại sinh thông qua MHC lớp II |
|||||
|
|
|
||||
 Hình 3
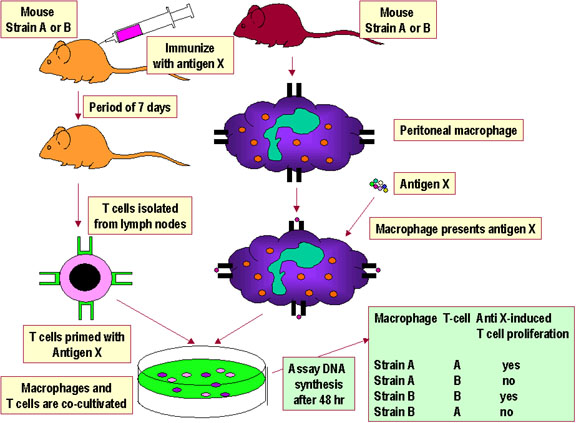
Hình 3Tương tác Th/APC được giới hạn trong MHC tự thân. |
|||||
 Hình 4
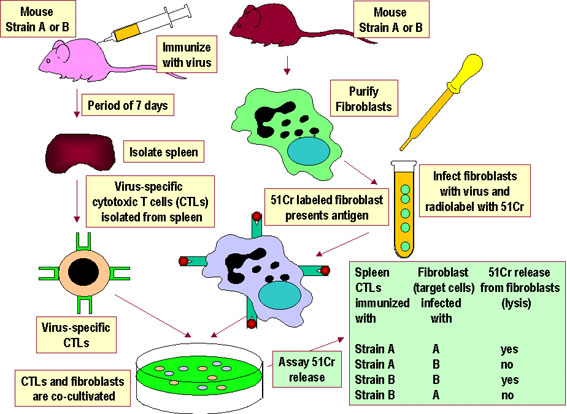
Hình 4Các CTL đặc hiệu virút từ chuột dòng A hay dòng B chỉ làm ly giải tế bào đích cùng dòng bị nhiễm một loại virút đặc hiệu. Các CTL không ly giải các tế bào đích không bị nhiễm virut và không đồng gen. Các nghiên cứu đã chỉ ra rằng các tế bào CTL và tế bào đích phải từ loài động vật có chung các allen MHC lớp I để các tế bào đích trình diện kháng nguyên của virút cho CTL. |
GIỚI HẠN MHC TỰ THÂN Để một tế bào T có thể nhận biết và đáp ứng với một kháng nguyên protein ngoại lai, trước tiên nó phải nhận ra MHC của các tế bào trình diện kháng nguyên là MHC của bản thân. Điều đó được gọi là giới hạn MHC tự thân. Các tế bào T hỗ trợ nhận biết kháng nguyên được trình diện bởi MHC tự thân lớp II, trong khi đó tế bào T gây độc tế bào lại nhận ra kháng nguyên trình diện bởi MHC lớp I. Quá trình mà các tế bào T bị giới hạn nhận biết các phân tử MHC tự thân diễn ra ở trong tuyến ức. Bằng thực nghiệm, người ta chứng minh giới
hạn MHC tự thân đối với tương tác tế bào T-APC và đối với tương tác tế bào T
gây độc-MHC được thể hiện trong Hình 3 và 4, một cách tương ứng. |
||||
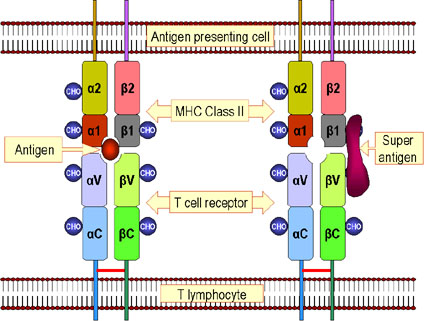 Hình 5
Hình 5Sự khác nhau giữa kháng nguyên và siêu kháng nguyên. Các peptid kháng nguyên được xử lý bên trong tế bào và được trình diện trên bề mặt tế bào ở dạng kết hợp với các phân tử MHC lớp II. Sau đó chúng hoạt hóa thụ thể tế bào T trên bề mặt của lympho T. Siêu kháng không được xử lý nhưng liên kết với các protein MHC lớp II và chuỗi beta V của thụ thể tế bào T. Một siêu kháng nguyên hoạt hóa một lớp tế bào T riêng biệt có biểu lộ chuỗi beta V nhất định. Lưu ý: Trong trường hợp
tương tác MHC II-TCR với một peptid được xử lý thông thường, sự nhận
biết các peptid trên các phân tử MHC đòi hỏi có các phân đoạn alpha V,
alpha J, beta V, beta D và beta J của TCR này. Tương tác như vậy xảy ra
ở tần suất thấp. Trong trường hợp tương tác MHC II-TCR với một siêu
kháng nguyên chưa qua xử lý, chỉ có một vùng beta V được nhận biết. Rõ
ràng điều này sẽ xảy ra ở một tần suất cao hơn nhiều. |
TẾ BÀO TRÌNH DIỆN KHÁNG NGUYÊN Có ba loại tế bào trình diện kháng nguyên chính, đó là tế
bào có tua (đuôi gai), các đại thực bào và tế bào B. Ngoài ra, một số các tế
bào khác có biểu lộ các phân tử MHC lớp II (ví dụ, các tế bào biểu mô tuyến
ức) có thể hoạt động như các tế bào trình diện kháng nguyên trong một số
trường hợp. Tế bào có tua (đuôi gai) được tìm thấy ở da và các mô khác,
chúng ăn kháng nguyên bởi ẩm bào và vận chuyển đến các hạch bạch huyết và
lách. Tại hạch bạch huyết và lách các tế bào này phân bố chủ yếu trong vùng
tế bào T. Tế bào đuôi gai là các tế bào trình diện kháng nguyên hiệu quả
nhất và có thể trình diện kháng nguyên cho các tế bào T trinh tiết. Hơn nữa,
chúng có thể trình diện các kháng nguyên nội bào đã được gắn lên phân tử MHC
lớp I hoặc lớp II (trình diện chéo), mặc dù con đường chủ yếu để trình diện
kháng nguyên là qua phân tử MHC lớp II. Loại tế bào thứ hai trình diện kháng
nguyên là đại thực bào. Những tế bào này ăn kháng nguyên bằng cách thực bào
hoặc ẩm bào. Các đại thực bào trình diện kháng nguyên cho các tế bào T trinh
tiết không có hiệu quả, nhưng lại rất tốt trong việc hoạt hóa tế bào T nhớ.
Loại thứ ba của tế bào trình diện kháng nguyên là tế bào B. Các tế bào này
liên kết với các kháng nguyên thông qua kháng thể bề mặt của chúng và bắt
kháng nguyên theo cơ chế ẩm bào. Giống như các đại thực bào, các tế bào này
không có hiệu quả như tế bào đuôi gai trong trình diện kháng nguyên cho tế
bào T trinh tiết. Các tế bào B trình diện kháng nguyên rất hiệu quả cho tế
bào T nhớ, đặc biệt là khi nồng độ kháng nguyên thấp bởi vì kháng thể bề mặt
của các tế bào B liên kết kháng nguyên với một ái lực cao. |
||||
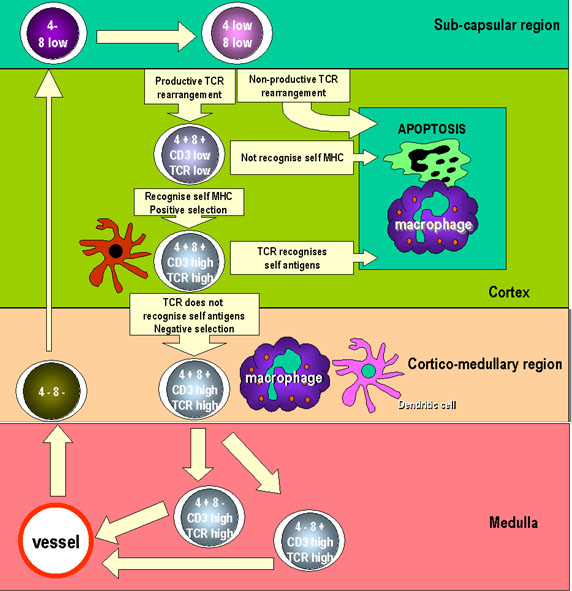 Hình 6
Hình 6Tiền tế bào T đi vào tuyến ức và phân chia thành lympho non có hình dạng lớn ở vùng dưới vỏ của tuyến ức. Các lympho non nhân lên và hình thành một bể chứa các tế bào biệt hóa. Ở đây các tế bào có dấu ấn CD8+ và CD4+ nhưng được biểu lộ thấp. Các gen TCR cũng được tái tổ hợp trong các tế bào này và các sản phẩm gen cũng được biểu lộ trên bề mặt tế bào ở mức độ thấp. Khi các tế bào trưởng thành, chúng di chuyển vào trong vùng vỏ, nơi chúng bám chặt vào các tế bào biểu mô vùng vỏ, đó là các tế bào dài và có nhiều tua tạo một diện tích bề mặt lớn để tương tác với các tế bào khác. Các TCR trên bề mặt của thymo bào tương tác với các phân tử MHC trên các tế bào biểu mô, dẫn đến chọn lọc dương tính. Các tế bào không được chọn sẽ bị chết theo chương trình và bị thực bào bởi các đại thực bào. Khi các thymo bào di chuyển sâu hơn vào vỏ tuyến ức thì sự biểu lộ CD3, CD4 CD8, và TCR tăng lên. Các TCR tự phản ứng sẽ mất đi vì tương tác với các tế bào đuôi gai và các đại thực bào. Điều này dẫn đến chọn lọc âm tính. Các tế bào biểu lộ CD4 hoặc CD8 xuất hiện và di chuyển ra ngoại vi bằng các ống đặc biệt đến vùng vỏ-tủy. |
ĐÀO TẠO Ở TUYẾN ỨC Cả hai loại tế bào Th và Tc bị gới hạn bởi MHC tự thân. Ngoài ra, tế bào T thường không nhận ra kháng nguyên tự thân. Làm thế nào để tạo ra các tế bào T giới hạn MHC tự thân và tại sao các tế bào T tự phản ứng không được hình thành? Sự tái tổ hợp ngẫu nhiên của VDJ trong tế bào T dẫn đến hình thành những tế bào T có khả năng nhận biết MHC không tự thân và một số tế bào T nhận biết MHC tự thân. Đó là vai trò của tuyến ức để đảm bảo rằng chỉ những tế bào T đi ra ngoại vi là loại có MHC giới hạn tự thân và không thể phản ứng với kháng nguyên bản thân. Các tế bào T có chức năng ở ngoại vi có để nhận biết kháng nguyên ngoại lai đã gắn với MHC tự thân, bởi vì các tế bào APC hoặc tế bào đích trình diện kháng nguyên ngoại lai liên kết với MHC tự thân. Tuy nhiên, một cá thể không cần tế bào T có chức năng ở ngoại vi để nhận biết kháng nguyên (tự thân hoặc ngoại lai) liên kết với MHC ngoại lai. Đặc biệt một cá thể không muốn chức năng tế bào T ở ngoại vi có thể nhận ra kháng nguyên tự thân liên kết với MHC tự thân vì chúng có thể làm tổn thương đến các mô bình thường, khỏe mạnh. Nhờ kết quả của sự kiện tái tổ hợp ngẫu nhiên VDJ xảy ra trong các tế bào T chưa trưởng thành ở trong tuyến ức mà các TCR được sản xuất có đầy đủ tính đặc hiệu. Các quá trình trong tuyến ức sẽ xác định sự đặc hiệu TCR được giữ lại. Có hai bước tuần tự được thể hiện trong Hình 6. Trước tiên, các tế bào T mà có khả năng liên kết với các phân tử MHC tự thân được biểu lộ bởi tế bào biểu mô vỏ tuyến ức thì được giữ lại. Đó là chọn lọc dương. Những tế bào nào không liên kết MHC tự thân thì sẽ bị chết theo chương trình. Do đó, tế bào T có một TCR nhận biết MHC tự thân tồn tại. Tiếp theo, những tế bào T mà có khả năng liên kết với các phân tử MHC tự thân kết hợp với các phân tử tự thân được biểu lộ bởi các tế bào biểu mô tuyến ức, tế bào đuôi gai và các đại thực bào thì sẽ bị tiêu diệt. Điều này được gọi là chọn lọc âm. Những tế bào không liên kết MHC tự thân thì được giữ lại. Sau hai bước đó, tế bào T có một TCR nhận biết MHC tự thân và kháng nguyên ngoại lai được sống sót. Mỗi tế bào T còn sống sau lựa chọn âm và dương trong tuyến ức và được đưa ra ngoại vi vẫn giữ được thụ thể tế bào T đặc hiệu của mình. Khi chọn lọc âm và dương diễn ra trong tuyến
ức, các tế bào T còn non cũng biểu lộ kháng nguyên CD4 hoặc CD8 trên bề mặt
của chúng. Ban đầu các tế bào tiền T đó đi vào tuyến ức là CD4-CD8-. Trong
tuyến ức nó sẽ trở thành CD4+CD8+ và khi chọn lọc âm và dương xảy ra, một tế
bào sẽ trở thành CD4+ hay CD8+. Việc trở thành tế bào CD4+ hay CD8+ phụ
thuộc vào loại phân tử MHC nào có mặt trên tế bào. Nếu một tế bào CD4+CD8+
được trình diện bởi phân tử MHC lớp I, thì CD4 giảm biểu lộ và nó trở thành
một tế bào CD8+. Nếu một tế bào được trình diện bởi các phân tử MHC lớp II,
thì CD8 giảm biểu lộ và trở thành một tế bào CD4+ (Hình 7). Chọn lọc âm và dương ở tuyến ức không phải là
quá trình có hiệu quả 100%. Hơn nữa, không phải tất cả các tự kháng nguyên
có thể được biểu lộ trong tuyến ức và dẫn đến một số tế bào T tự phản ứng có
thể ra ngoại vi. Như vậy, có những cơ chế bổ sung được tạo ra để loại bỏ tế
bào T tự phản ứng ở ngoại vi và điều đó sẽ được thảo luận trong chương dung
thứ. |
||||
|
|||||
|
|
|||||
|
|||||
|
|
This page last changed on
Monday, September 18, 2017
|
||||