|
x |
x |
|
 |
 |
|
BỆNH NHIỄM TRÙNG |
VI KHUẨN HỌC |
MIỄN DỊCH HỌC |
NẤM HỌC |
KÝ SINH TRÙNG HỌC |
VIRÚT HỌC |
|
ENGLISH |
MIỄN DỊCH HỌC – CHƯƠNG HAI
BỔ THỂ
Gene Mayer, Ph.D
Emertius Professor of Pathology, Microbiology and Immunology
University of South Carolina
Biên dịch: Nguyễn Văn Đô, MD.,
PhD.,
Bộ môn Sinh lý bệnh-Miễn dịch,
Trường Đại học Y Hà Nội,
Hà Nội, Việt Nam
|
|
TURKISH |
|
FRANCAIS |
|
SHQIP |
|
ESPANOL |
|
PORTUGUES |
|
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|
|

 |
|
Logo image © Jeffrey
Nelson, Rush University, Chicago, Illinois and
The MicrobeLibrary |
|
|
|
MỤC TIÊU HỌC TẬP
Hiểu được các con đường hoạt hóa bổ thể khác nhau
Biết được các cơ chế hoạt hóa bổ thể do enzym và không do emzym
Biết được các đặc điểm sinh học của các sản phẩm hoạt hóa bổ thể
Biết được ý nghĩa của hệ thống bổ thể trong đề kháng của cơ thể, viêm và
tự tổn thương của cơ thể
Hiểu được các cơ chế điều hòa hoạt hóa bổ thể và các sản phẩm của chúng
 Jules Bordet (1870-1961), Khám phá ra bổ thể
Jules Bordet (1870-1961), Khám phá ra bổ thể
National Library of Medicine
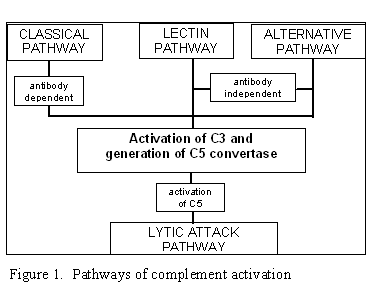
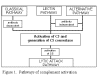 Hình 1. Các con đường hoạt hóa bổ thể
Hình 1. Các con đường hoạt hóa bổ thể |
CHỨC NĂNG BỔ THỂ
Trong lịch sử, từ bổ thể (viết tắt là C) được sử dụng để chỉ một
thành phần trong huyết thanh không bền với nhiệt có thể ly giải vi khuẩn (hoạt
tính của C bị tiêu hủy (bất hoạt) bằng cách đun nóng huyết thanh ở 560C trong 30
phút). Tuy nhiên, bổ thể hiện đang được biết là góp phần vào sự bảo vệ của cơ
thể chủ bằng những con đường khác nhau. Bổ thể có thể opsonin hóa vi khuẩn để
làm tăng cường sự thực bào, tập trung và hoạt hóa các tế bào khác nhau bao gồm
các tế bào bạch cầu đa nhân (PMN) và các đại thực bào, tham gia vào sự điều hòa
các đáp ứng của kháng thể và hỗ trợ trong việc dọn dẹp phức hợp miễn dịch và các
tế bào chết theo chương trình. Bổ thể cũng có thể có tác động có hại cho cơ thể
chủ như góp phần vào tình trạng viêm, làm tổn thương mô và có thể gây ra sốc
phản vệ.
Bổ thể bao gồm hơn 20 protein huyết thanh khác
nhau (xem Bảng 1) được sản xuất bởi một loạt các tế bào bao gồm: tế bào gan, đại
thực bào và tế bào biểu mô ruột. Một số protein bổ thể liên kết với globulin
miễn dịch hoặc các thành phần của màng tế bào. Nhiều thành phần khác ở dạng tiền
enzym, khi hoạt hóa sẽ phân cắt một hoặc nhiều protein bổ thể khác. Sau khi cắt
một số các protein của bổ thể thì tạo ra các mảnh có khả năng hoạt hóa tế bào,
tăng tính thấm thành mạch hoặc opsonin hóa vi khuẩn.
|
Bảng 1. Các protein của bổ
thể
|
|
Đường cổ điển |
Đường Lectin |
Đường cạnh |
Đường ly giải |
|
Protein hoạt hóa:
C1qrs, C2, C3, C4
Protein điều hòa:
C1-INH, C4-BP
|
Protein liên kết manose (MBP), Manose liên kết với serine
của protease (MASP, MASP2) |
C3, yếu tố B & D*, Properdin (P)
Yếu tố I* và H, yếu tố tăng phân hủy (DAF), thụ thể 1của bổ thể (CR1),
etc.
|
C5, C6, C7, C8, C9
Protein S |
|
Các thành phần được gạch chân có hoạt tính enzym
khi hoạt hóa.
Những thành phần được đánh dấu với một dấu sao có hoạt tính enzym ở dạng tự
nhiên của chúng
|
|
| |
CÁC CON ĐƯỜNG HOẠT HÓA BỔ THỂ
Hoạt hóa bổ thể có thể được chia thành bốn con
đường (Hình 1): con đường cổ điển, con đường lectin, con đường cạnh và con đường
tấn công màng (hoặc ly giải). Cả hai con đường cổ điển và đường cạnh dẫn đến
hoạt hóa convertase C5 và kết quả là tạo ra C5b đó là yếu tố cần thiết để hoạt
hóa con đường tấn công màng.
|
|
MOVIE
Complement Activation and Biological Functions
High Resolution
Quicktime
Low Resolution Quicktime
© Scott R. Barnum, University of Alabama, Birmingham, Ala., USA and
The
MicrobeLibrary
|
|
CGAP
More
detailed complement pathways from CGAP/Biocarta |
Con đường
cổ điển (Hình 2)
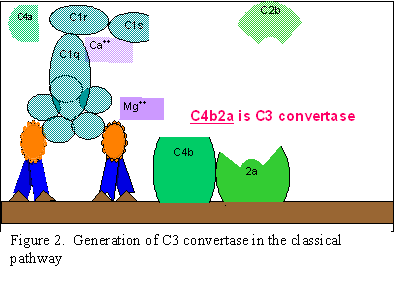
Hoạt hóa C1
C1, một protein
đa tiểu đơn vị có ba protein khác nhau (C1q, C1r và C1s), liên kết với
vùng Fc của các phân tử kháng thể IgG và IgM đã kết hợp với kháng nguyên.
Sự liên kết đó của C1 không xảy ra nếu kháng thể không kết hợp với kháng
nguyên và không có mặt của các ion Canxi và Magiê. (lưu ý, trong một số
trường hợp C1 có thể liên kết với globulin miễn dịch vón tụ [ví dụ như
vón tụ IgG] hay trên bề mặt tác nhân gây bệnh nhất định khi không có
kháng thể). Sự liên kết của C1 với kháng thể thông qua C1q và C1q phải
có ít nhất hai liên kết với phân tử kháng thể trước khi nó được cố định
vững chắc. Sự liên kết của C1q làm cho hoạt hóa C1r rồi sau đó hoạt hóa
C1s. Kết quả là hình thành "C1qrs” hoạt hóa, một loại enzym mà sẽ cắt C4
thành hai mảnh C4a và C4b.
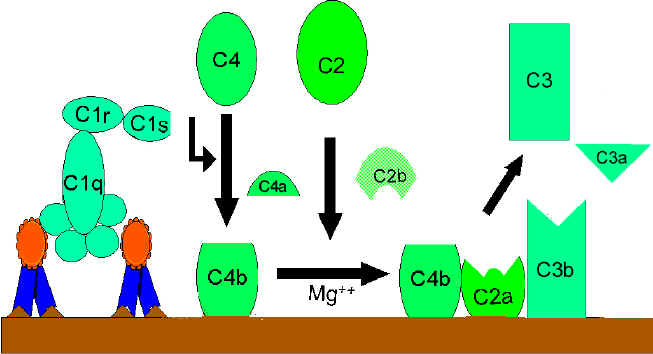
Hoạt hóa C4 và C2 (tạo ra convertase C3)
Mảnh C4b liên
kết với màng và mảnh C4a được giải phóng ra môi trường xung quanh.
"C1qrs" hoạt hóa cũng cắt C2 thành C2a và C2b. C2a gắn lên màng tế bào
cùng với C4b còn C2b được giải phóng ra môi trường. Phức hợp C4bC2a là
một convertase C3, enzym này sẽ cắt C3 thành C3a và C3b.
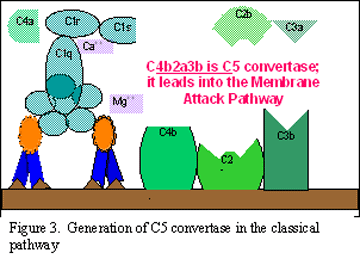
Hoạt hóa C3 (tạo ra convertase C5)
C3b gắn vào màng
tế bào trong phức hợp với C4b và C2a, còn C3a được đưa ra môi trường
xung quanh. Kết quả là tạo ra C4bC2aC3b, là một convertase C5. Sự hình
thành convertase C5 là phần cuối của con đường cổ điển.
Một số sản phẩm
của con đường cổ điển có những hoạt tính sinh học mạnh góp phần vào bảo
vệ cơ thể chủ. Các sản phẩm này cũng có thể tác động có hại nếu được sản
xuất mà không có kiểm soát. Bảng 2 tóm tắt các hoạt tính sinh học của
các thành phần trong con đường cổ điển.
|
Bảng 2. Hoạt tính sinh
học của các thành phần con đường cố điển |
|
Thành phần |
Hoạt tính sinh học |
| C2b |
Prokinin; phân cắt
bởi plasmin tạo ra nhiều kinin, dẫn đến phù |
| C3a |
Phản vệ; có thể
hoạt hóa các tế bào mast và tế bào bạch cầu ái kiềm để giải phóng
hạt dẫn đến tăng tính thấm thành mạch và co thắt cơ trơn, có thể dẫn
đến sốc phản vệ. |
| C3b |
Opsonin; thúc đẩy thực bào bằng cách gắn vào thụ thể
của bổ thể Hoạt hóa tế bào thực bào |
| C4a |
Phản vệ; (yếu
hơn C3a) |
| C4b |
Opsonin; thúc đẩy
thực bào bằng cách gắn vào thụ thể của bổ thể |
Nếu con đường cổ điển không được điều hòa thì C2b, C3a, và C4a vẫn được
tiếp tục sản xuất.Vì vậy, phải có một số cách để điều hòa hoạt động của
con đường cổ điển. Bảng 3 tóm tắt các cách thức mà con đường cổ điển
được điều hòa.
|
Bảng 3. Điều hòa con
đường cổ điển |
| Thành phần |
Điều hòa |
| Tất cả |
C1-INH; C1r và C1s
tách ra từ C1q |
| C3a |
Bất hoạt C3a
(C3a-INA;Carboxypeptidase B); Bất hoạt C3a |
| C3b |
Yếu tố H và I; Yếu
tố H tạo điều kiện cho sự tiêu hủy C3b bởi yếu tố Iactor H
facilitates the degradation of C3b by Factor I |
| C4a |
C3-INA |
| C4b |
Protein in gắn C4
(C4-BP) và yếu tố I; C4-BP tạo điều kiện cho tiêu hủy C4b bởi yếu tố
I; C4-BP cũng ngăn ngừa kết hợp của C2a với C4b do đó ngăn chặn sự
hình thành convertase C3. |
Tầm quan trọng của INH-C1
trong điều hòa con đường cổ điển được thể hiện bằng kết quả của sự thiếu
hụt chất ức chế này. Sự suy giảm INH-C1 liên quan đến sự phát triển của
phù mạch di truyền
|
 Hình 2.
Hình 2.
A. Tạo convertase C3 ở con đường cổ điển
 B. Tạo convertase C5 ở con đường cổ điển
B. Tạo convertase C5 ở con đường cổ điển

C. Hoạt hóa C3 theo con đường cổ điển
|
 Hình 3.
Hình 3.
Con đường khởi đầu bởi lectin |
Con đường lectin
Con đường lectin (Hình 3) rất giống với con đường cổ
điển. Nó được khởi đầu bằng cách lectin liên kết với mannose (MBL) nằm ở
phân tử polysaccharid của bề mặt vi khuẩn. Sự kết hợp của MBL vào một
mầm bệnh dẫn đến hình thành phức hợp MBL với hai protease serine là
MASP-1 và MASP-2 (MBL liên kết với hai protease ở vị trí serine). MASP-1
và MASP-2 cũng tương tự như C1r và C1s một cách tương ứng và MBL cũng
tương tự như C1q. Sự hình thành phức hợp ba phân tử MBL/MASP-1/MASP-2 sẽ
hoạt hóa các phân tử MASP và sau đó cắt C4 thành C4a và C4b. Các mảnh
C4b liên kết với màng tế bào đích và mảnh C4a được đưa vào môi trường
xung quanh. Hoạt hóa MASP cũng cắt C2 thành C2a và C2b. C2a gắn lên màng
tế bào cùng với C4b, còn C2b được đưa vào môi trường. Phức hợp C4bC2a là
enzym convertase C3, nó sẽ cắt C3 thành C3a và C3b. C3b gắn vào màng tế
bào cùng với C4b và C2a, còn C3a được đưa vào môi trường. Kết quả là
hình thành phức hợp C4bC2aC3b, đây là enzym convertase C5. Sự hình thành
convertase C5 là phần cuối của con đường lectin.
Các hoạt tính sinh học và các
protein điều hòa của con đường
lectin đều giống con đường cổ điển.
|
 Hình
4. Hình
4.
Hoạt hóa C3 tự phát |
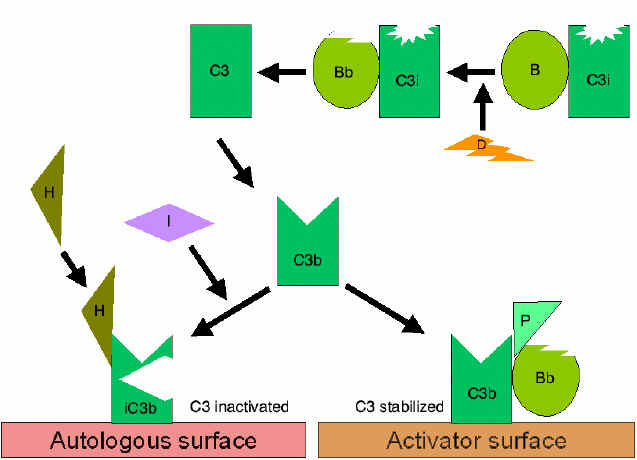
Con đường cạnh
Con đường cạnh bắt đầu với sự hoạt hóa C3 và cần có
yếu tố B, D và cation Mg++, chúng đều có mặt trong huyết thanh người
bình thường.
Khuếch đại vòng lặp tạo ra C3b
(Hình 4)
Trong huyết thanh, sự thủy phân C3 để tạo ra C3i
được duy trì một cách tự phát ở một mức độ thấp. Yếu tố B liên kết
với C3i và trở thành phức hợp nhạy cảm cho yếu tố D để nó cắt yếu tố
B thành Bb. Các phức hợp C3iBb có vai trò như một convertase C3 và
sẽ cắt C3 thành C3a và C3b. Một khi C3b được hình thành, yếu tố B sẽ
liên kết với nó và trở nên nhạy cảm cho yếu tố D phân cắt yếu tố B.
Phức hợp C3bBb là một convertase C3 sẽ tiếp tục tạo ra nhiều C3b, vì
thế sản phẩm C3b được khuếch đại. Nếu quá trình này diễn ra liên tục
mà không được kiểm soát, thì C3 trong huyết thanh sẽ bị tiêu thụ hết.
Do đó, việc sản xuất C3b tự phát phải được kiểm soát một cách chặt
chẽ.
|

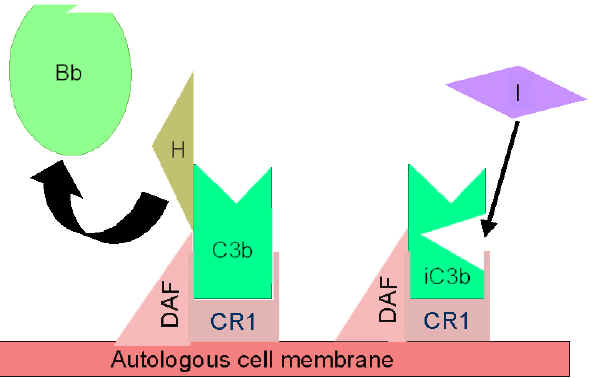
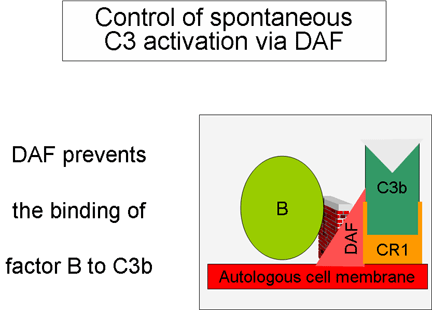
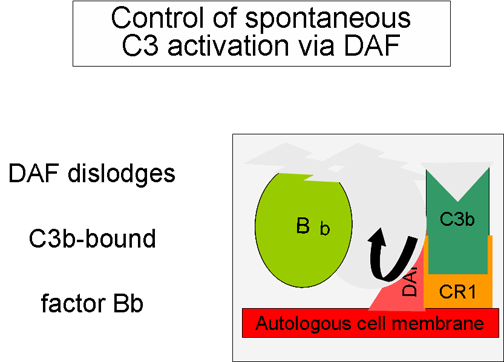
 Hình 5. Điều hòa C3 hoạt hóa bởi DAF
Hình 5. Điều hòa C3 hoạt hóa bởi DAF
 Hình
6. Hình
6.
Điều hòa C3 hoạt hóa bởi Cr1
 Hình
7. Hình
7.
Ổn định convertase C3
 Hình 8.
Hình 8.
Convertase C5 của con đường cạnh trở nên ổn định |
Kiểm soát vòng lặp khuếch đại
(Hình 5 và 6)
Khi
C3b, sản phẩm được tạo một cách tự nhiên, liên kết với màng tế bào tự
thân và tương tác với DAF (yếu tố thúc đẩy sự phân rã) sẽ ngăn chặn sự
kết hợp của yếu tố B với C3b dẫn đến ngăn ngừa sự hình thành convertase
C3. Ngoài ra, DAF tăng tốc độ phân ly của Bb từ phức hợp C3bBb (convertase
C3), do đó ngăn cản sự sản xuất thêm C3b. Một số tế bào có thụ thể 1 với
bổ thể (CR1). Sự liên kết của C3b với CR1 tạo điều kiện phân hủy enzym
của C3b bởi yếu tố I. Ngoài ra, liên kết của convertase C3 (C3bBb) với
CR1 cũng phân ly Bb từ phức hợp. Như vậy, trong các tế bào có thụ thể
với bổ thể, CR1 cũng đóng một vai trò trong việc kiểm soát vòng lặp
khuếch đại. Cuối cùng, yếu tố H có thể liên kết với C3b đã gắn vào một
tế bào hoặc ở pha lỏng làm cho sự tiêu hủy enzym của C3b bởi yếu tố I.
Như vậy, vòng lặp khuếch đại được kiểm soát bằng cách ngăn chặn sự hình
thành convertase C3, phân ly convertase C3, hoặc tiêu hóa C3b bằng enzym.
Tầm quan trọng của việc kiểm soát vòng lặp khuếch đại được minh họa ở
những bệnh nhân thiếu sót di truyền của yếu tố H hoặc I. Những bệnh nhân
này thiếu C3 và tăng nhạy cảm với một số nhiễm trùng nhất định.
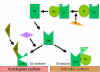
Ổn định của C convertase bởi các bề mặt chất hoạt hóa (bảo vệ)
(Hình 7)
Khi bám vào một yếu tố kích thích phù hợp của con đường cạnh, C3b sẽ
liên kết với yếu tố B và với sự phân cắt enzym của yếu tố D thì
convertase C3 (C3bBb) được hình thành. Tuy nhiên, C3b không có khả năng
phân hủy bởi yếu tố I và convertase C3 không bị tiêu đi nhanh chóng, vì
nó được ổn định bởi bề mặt chất hoạt hóa. Phức hợp được ổn định hơn nữa
nếu properdin bám vào C3bBb. Các chất hoạt hóa của con đường cạnh là các
thành phần trên bề mặt của tác nhân gây bệnh và bao gồm: LPS của vi
khuẩn gram âm và thành tế bào của một số vi khuẩn và men bia. Vì vậy,
khi C3b liên kết với một bề mặt hoạt hóa, các convertase C3 hình thành
sẽ được ổn định và tiếp tục tạo thêm C3a và C3b từ sự phân cắt C3.
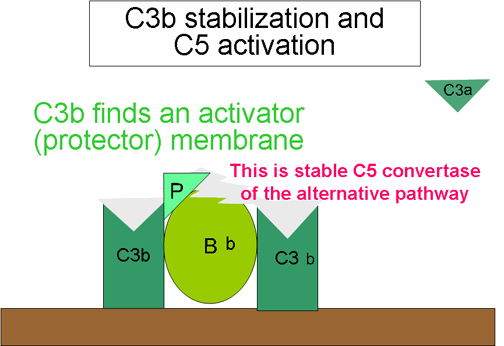
Sự hình thành
convertase C5 (Hình 10)
Một số C3b được tạo ra bởi các convertase C3 ổn định
trên bề mặt chất hoạt hóa với phức hợp C3bBb để tạo thành một phức
hợp C3bBbC3b. Phức hợp này gọi là convertase C5 và cũng là chặng
đường cuối cùng của con đường cạnh. Con đường này có thể được hoạt
hóa bởi nhiều vi khuẩn gram âm (tiêu biểu là Neisseria meningitidis
và N. gonorrhoea), một số vi khuẩn gram dương, virut và ký sinh
trùng, và hậu quả là các vi sinh vật bị ly giải. Như vậy, hoạt hóa
bổ thể theo con đường cạnh giúp cho cơ thể một phương tiện bảo vệ
chống lại các tác nhân gây bệnh nhất định trước khi một phản ứng
kháng thể xảy ra. Sự thiếu hụt C3 làm gia tăng tính nhạy cảm với
những vi sinh vật. Con đường cạnh có thể là con đường nguyên thủy,
còn con đường cổ điển và lectin có thể phát triển từ con đường cạnh.
|
| |
|
| |
Hãy nhớ rằng con đường cạnh cung cấp một
phương tiện đề kháng không đặc hiệu chống lại nhiễm trùng mà không có sự
tham gia của các kháng thể và do đó tạo ra hàng rào bảo vệ đầu tiên chống
lại một số tác nhân gây bệnh.
Nhiều vi khuẩn gram âm và một số vi khuẩn gram dương, một số vi rút, các ký
sinh trùng, dị hồng cầu, globulin miễn dịch vón tụ (đặc biệt, IgA) và một số
protein khác (ví dụ như protease, các sản phẩm đông máu) có thể hoạt hóa con
đường cạnh. Một protein là yếu tố nọc độc rắn hổ mang (CVF) đã được nghiên
cứu một cách tích cực về khả năng hoạt hóa con đường này của nó.
|
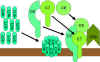 Hình 9.
Hình 9.
Con đường ly giải |
Con đường tấn công màng (ly giải) (Hình
9)
Convertase C5 được tạo ra từ con đường cổ điển (C4b2a3b),
lectin (C4b2a3b) hoặc con đường cạnh (C3bBb3b) sẽ cắt C5 thành C5a và C5b.
C5a ở lại pha lỏng, còn C5b nhanh chóng liên kết với C6 và C7, rồi sau đó
chúng bám lên màng tế bào đích. Tiếp theo, C8 liên kết với chúng và cuối
cùng là nhiều mảnh của phân tử C9 gắn vào phức hợp đó. Các mảnh của C9 tạo
thành một lỗ thủng xuyên màng, các thành phần bên trong tế bào bị rò rỉ và
ly giải diễn ra. Ly giải không phải là một quá trình enzym, nó được cho là
do các tổn thương cơ học màng tế bào. Phức hợp bao gồm C5bC6C7C8C9 được gọi
là phức hợp tấn công màng (MAC).
C5a được sinh ra từ con đường ly giải có một số hoạt tính
sinh học mạnh. Đây là loại gây phản vệ mạnh nhất. Ngoài ra, C5a là một yếu
tố hóa hướng động bạch cầu trung tính, tăng hô hấp tế bào và kích thích các
đại thực bào sản xuất cytokin gây viêm. Hoạt tính của C5a được bất hoạt bởi
carboxypeptidase B (C3-INA).
Một số phức hợp C5b67 có thể được bong ra khỏi màng và đi
vào pha lỏng. Nếu điều này xảy ra, nó có thể liên kết với các tế bào khác
gần đó và dẫn đến ly giải chúng. Sự tổn thương các tế bào bên cạnh được ngăn
chặn bằng Protein S (vitronectin). Protein S liên kết với C5b67 hòa tan và
ngăn không cho chúng bám vào các tế bào khác.
|
 Hình
10. Điểu hòa C1rs (convertase C4) bởi C1-INH Hình
10. Điểu hòa C1rs (convertase C4) bởi C1-INH |
CÁC SẢN PHẨM CỦA HOẠT HÓA BỔ THỂ CÓ HOẠT TÍNH SINH
HỌC
Hoạt hóa bổ thể tạo ra một số sản phẩm có hoạt tính sinh học
góp phần vào sự bảo vệ cơ thể, sốc phản vệ và viêm.
Sản xuất kinin
C2b được tạo ra khi hoạt hóa con đường cổ điển là một tiền
kinin có hoạt tính sinh học theo sau là sự biến đổi enzym bởi plasmin.
C1-INH ngăn ngừa việc tạo ra quá nhiều C2b bằng cách hạn chế hoạt hóa C2,
C1-INH còn được gọi là serpin có tác dụng tách C1rs từ phức hợp C1qrs (Hình
10). Sự thiếu hụt di truyền của C1-INH làm cho C2b được tạo ra quá nhiều là
nguyên nhân của bệnh phù mạch di truyền. Tình trạng này có thể được điều trị
bằng Danazol để thúc đẩy sản xuất C1-INH hoặc với acid caproic amin-ε để làm
giảm hoạt tính của plasmin.
|
|
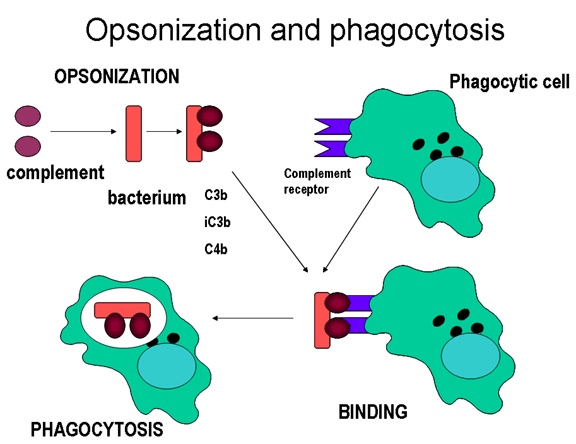
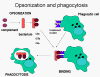 Hình 11.
Hình 11.
Protein bổ thể gắn lên bề mặt vi sinh vật và làm tăng thực bào thông qua
thụ thể của bổ thể.
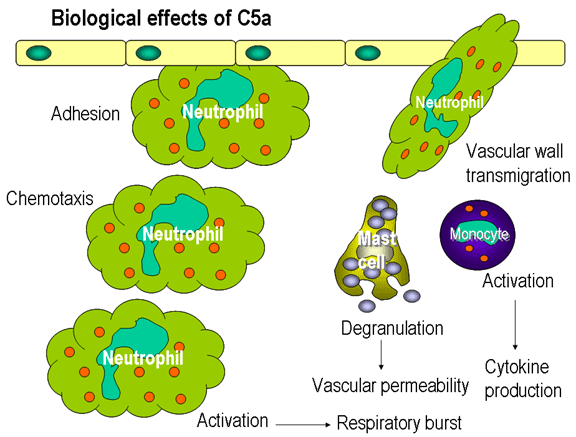
 Hình 12.
Hình 12.
Hoạt tính sinh học của C5a
|
Chất gây sốc phản vệ
C4a, C3a và C5a (theo thứ tự tăng dần của hoạt
tính) là các chất gây phản vệ, chúng làm giải phóng hạt và co thắt cơ trơn.
Tác dụng không mong muốn của các peptid này được kiểm soát bởi
carboxypeptidase B (C3a-INA).
Các yếu tố hóa hướng động
Cả C5a và MAC (C5b67) là các chất hóa hướng động.
C5a cũng có khả năng hoạt hóa bạch cầu trung tính, bạch cầu ái kiềm và các
đại thực bào và gây ra cảm ứng các phân tử bám dính vào các tế bào nội mạc
mạch máu (Hình 12).
Opsonin
Các mảnh C3b và C4b đã bám lên bề mặt của vi sinh
vật sẽ kết hợp với thụ thể của bổ thể (CR1) nằm trên bề mặt các tế bào thực
bào và thúc đẩy thực bào (Hình 11).
Các sản phẩm khác của hoạt hóa bổ thể có hoạt tính sinh học
Các sản phẩm của C3 (iC3b, C3d và C3e) cũng gắn
vào các tế bào khác nhau bằng các thụ thể riêng biệt và điều hòa chức năng
của chúng.
Tóm lại, hệ thống bổ thể tham gia vào cả hai loại
đề kháng miễn dịch đặc hiệu và không đặc hiệu, đồng thời giải phóng một số sản
phẩm có ý nghĩa sinh học và bệnh lý (Bảng 4).
Nhiều suy giảm các thành phần bổ thể mang tính di
truyền đã được biết, nhưng thiếu hụt C3 là nghiêm trọng nhất và gây tử vong.
Thiếu hụt bổ thể cũng xảy ra trong các bệnh do phức hợp miễn dịch (ví dụ, SLE),
nhiễm vi khuẩn, virut và ký sinh trùng cấp tính và mạn tính.
|
|
|
|
|
|
|
|
|
Bảng 4. Hoạt tính của
các sản phẩm do hoạt hóa bổ thể và các yếu tố kiểm soát |
| Mảnh |
Hoạt tính |
Tác dụng |
Các yếu tố kiểm soát |
| C2a |
Prokinin, tích
tụ chất lỏng |
Phù |
C1-INH |
| C3a |
Giải phóng hạt
của các tế bào mast hay bạch cầu ái kiềm; tăng tính thấm thành
mạch, co thắt cơ trơn |
Phản vệ |
C3a-INA |
| C3b |
Opsonin, hoạt
hóa tế bào thực bào |
Thực bào |
Yếu tố H và I |
| C4a |
Giải phóng hạt
của các tế bào mast hay bạch cầu ái kiềm; tăng kính thấm thành
mạch, co thắt cơ trơn |
Phản vệ
(yếu nhất)
|
C3a-INA |
| C4b |
Opsonin |
Thực bào |
C4-BP và yếu tố
I |
| C5a |
Giải phóng hạt
của các tế bào mast hay bach cầu ái kiềm; tăng tính thấm thành
mạch, co thắt cơ trơn |
Phản vệ
(Mạnh nhất)
|
C3a-INA |
| Hóa hướng động,
tăng hô hấp tế bào, hoạt hóa thực bào, kích thích các cytokin
viêm |
Viêm |
| C5bC6C7 |
Hóa hướng động |
Viêm |
Protein S
(vitronectin) |
| Tấn công màng
tế bào |
Tổn thương mô |
|
|
Bạn đã học được
Các
protein của hệ thống bổ thể
Sự giống nhau và khác nhau của các con đường hoạt hóa bổ
thể
Ý nghĩa của các con đường khác nhau trong đáp ứng miễn
dịch đặc hiệu và không đặc hiệu
Vai trò của
các sản phẩm hoạt hóa bổ thể khác nhau trong khuếch đại miễn dịch đặc
hiệu và không đặc hiệu và viêm
|
|
Bảng 5. Thiếu hụt bổ
thể và bệnh |
| Con đường/yếu tố |
Bệnh |
Cơ chế bệnh sinh |
| Đường cổ điển |
|
| C1INH |
Phù mạch di
truyền |
Sản xuất quá
nhiều C2b (prokinin) |
| C1, C2, C4
|
Xu hướng SLE |
Opsonin hóa của
phức hợp miễn dịch giúp chúng hòa tan, sự thiếu hụt làm tăng
lượng kết tụ ở các mô và viêm |
| Con đường Lectin |
|
| MBL |
Nhạy cảm với
nhiễm vi khuẩn ở trẻ sơ sinh hoặc người bị ức chế miễn dịch |
Không có khả
năng khởi động con đường lectin |
| Con đường cạnh |
|
| Yếu tố B hoặc D |
Nhạy cảm với
nhiễm vi khuẩn sinh mủ |
Thiếu sự
opsonin hóa vi khuẩn |
| C3 |
Nhạy cảm với
nhiễm vi khuẩn |
Thiếu sự
opsonin hóa và không có khả năng sử dụng con đường tấn công màng |
| C5, C6, C7 C8,
và C9 |
Nhạy cảm với
nhiễm vi khuẩn Gram âm |
Inability to attack the outer
membrane of Gram-negative bacteria |
| Properdin (liên
kết NST X) |
Nhạy cảm viêm
màng não do não mô cầu |
Thiếu sự
opsonin hóa vi khuẩn |
| Yếu tố H hoặc
I |
Sự thiếu hụt C3
và nhạy cảm với nhiễm vi khuẩn |
Không kiểm soát
được hoạt hóa C3 ở con đường cạnh dẫn đến sự suy giảm C3 |
|
|
|
 Trở về phần Miễn dịch của Vi khuẩn học và Miễn dịch học
online
Trở về phần Miễn dịch của Vi khuẩn học và Miễn dịch học
online
This page last changed on
Sunday, August 13, 2017
Page maintained by
Richard Hunt
|
 Hình 2.
Hình 2.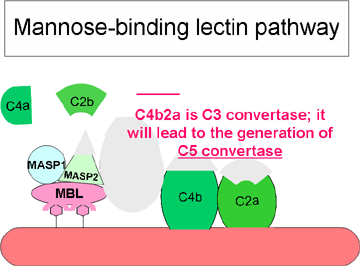 Hình 3.
Hình 3. 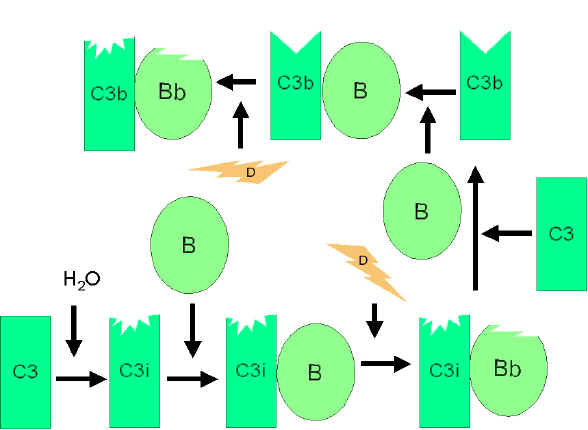 Hình
4.
Hình
4. 
 Hình 5. Điều hòa C3 hoạt hóa bởi DAF
Hình 5. Điều hòa C3 hoạt hóa bởi DAF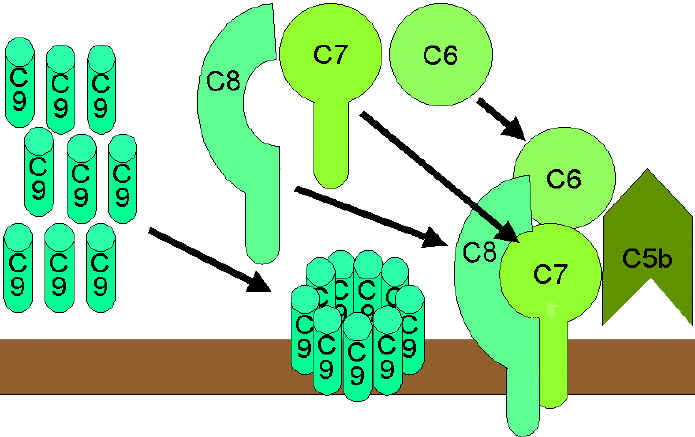 Hình 9.
Hình 9. 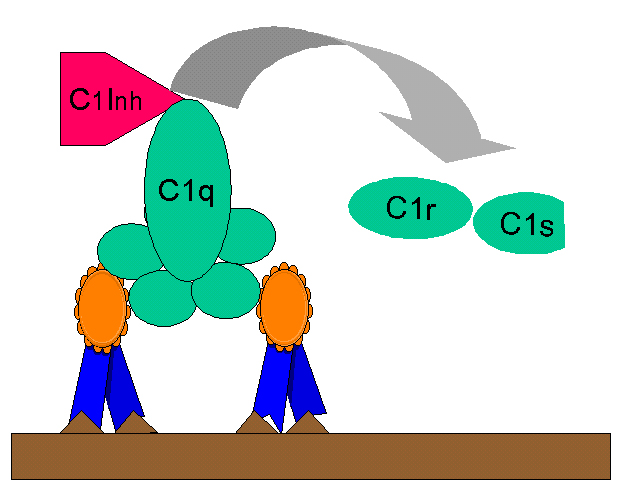 Hình
10. Điểu hòa C1rs (convertase C4) bởi C1-INH
Hình
10. Điểu hòa C1rs (convertase C4) bởi C1-INH